Información acerca de bacterias en nuestro blog

Otras bacterias (volumen V)

Gram+ con Alto Contenido en G+C: Actinobacterias

Gram+ con bajo contenido en G+C: Firmicutes. Molicutes

Otras Proteobacterias (II)

Rickettsias: otras Proteobacterias (I)

Proteobacterias quimioorganótrofas anaerobias facultativas

Proteobacterias quimiorganótrofas aerobias
Proteobacterias quimiolitótrofas y metanótrofas

Bacterias fotosintéticas

Aquifex y el grupo Deinococcus-Thermus
La célula procariota

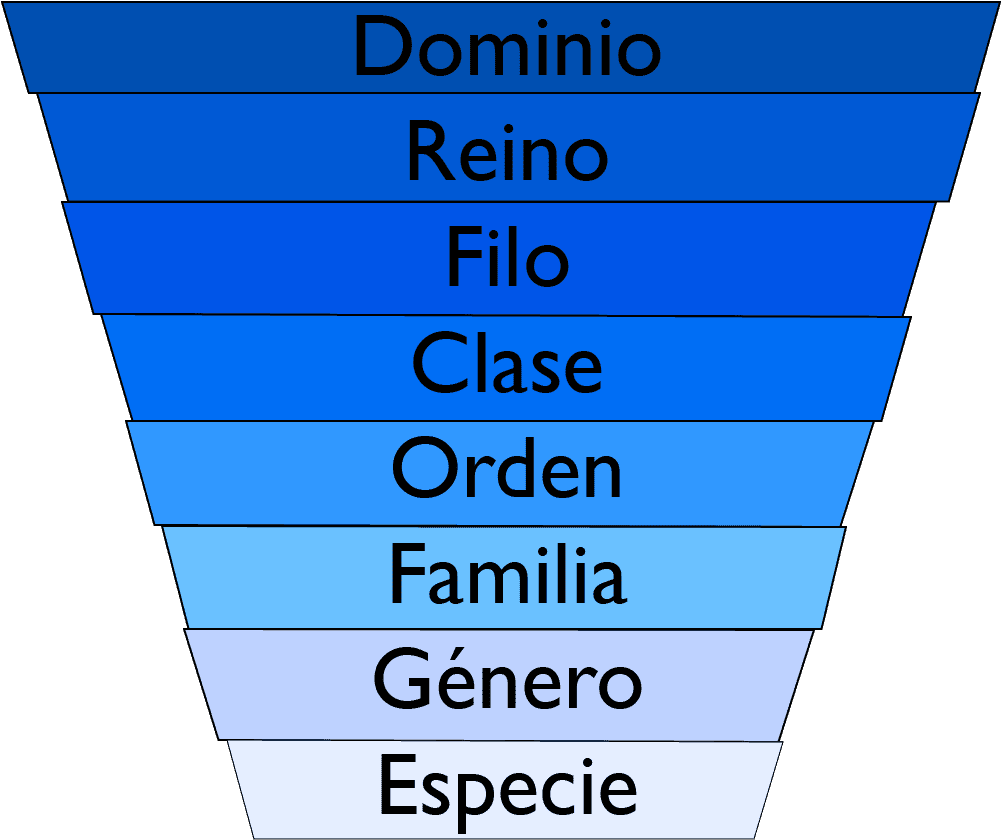
Clasificación de los seres vivos: taxonomía

El peptidoglicano: qué es, estructura y síntesis
Clasificación de las bacterias
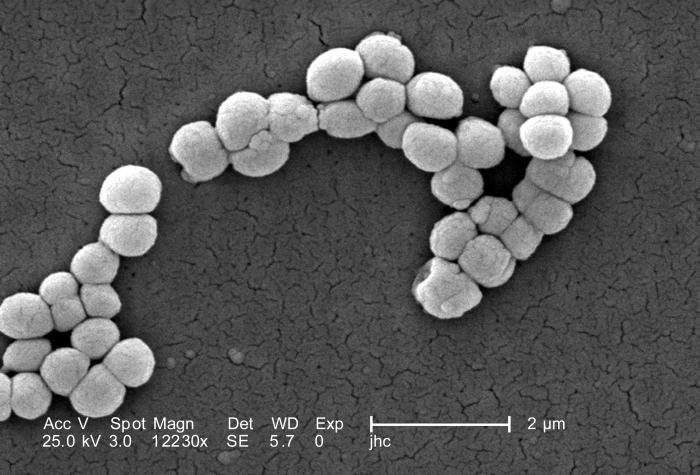
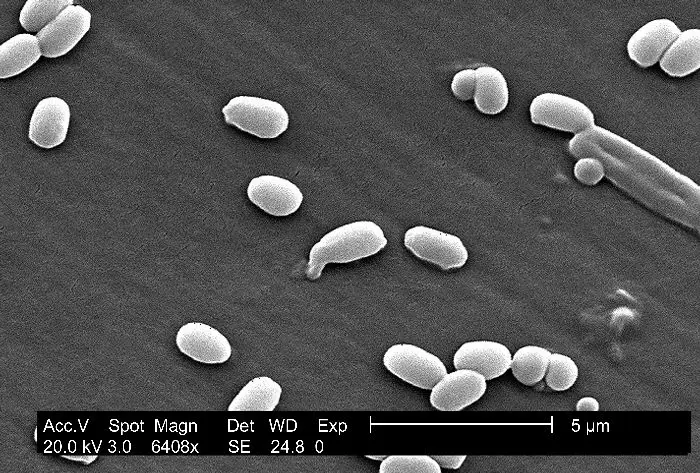
Bacterias: Caracteres generales