
Otras Proteobacterias (II)
bacterias, bacterias pedunculadas, bacterias reductoras de azufre, Bacterias reductoras de sulfato, Bdellovibrio, espirilos, grupo épsilon, Helicobacter, Helicobacter pylori, mixobacterias, mixobacterias deslizantes, proteobacterias, Vibrioides | 13 de mayo de 2021 | 20 min de lectura | ¿Prefieres oír el artículo? ¡Dale al ▶︎!
Cargando...
Configuración
En el último apartado de proteobacterias veremos a otras proteobacterias que no entran en ninguno de los grupos anteriores, a saber: proteobacterias con morfología inusual (espirilos, bacterias con vaina), pedunculadas, mixobacterias deslizantes, bacterias reductoras de sulfato del sulfato y azufre, y proteobacterias del grupo épsilon.
Índice de contenidos
Proteobacterias con morfología inusual: espirilos
Helicoidales
Dentro de las helicoidales entran Spirillum, Aquaspirillum, Oceanospirillum, Magnetospirillum o Azospirillum.
Todas cuentan con típica morfología helicoidal (forman más de una vuelta de hélice). Son móviles mediante flagelos polares generalmente en número superior a dos (pueden presentar penachos, flagelacion lofótrica) en ambos polos celulares. Tienden a ser microaerófilos. Son de gran tamaño en general. Hay especies acuáticas y terrestres. La mayoría son acuáticas e inofensivas como Spirillum (prototipo de este grupo).

Spirillum volutans
Es muy grande, de 1’6 x60 μm; se supone que podría ser el microorganismo que describió Leeuwenhoeck, el padre de la microbiología. Es microaerófilo, con flagelación lofótrica en ambos extremos. Tiene una acumulación de gránulos de volutina. Es un organismo acuático.
Aquaspirillum
Es aerobio o microaerófilo. Flagelación lofótrica en ambos extremos. Vive en agua dulce.
Oceanospirillum
Habita en agua salada, por lo que para poder cultivarlo debemos añadir NaCl al medio de cultivo. Es aerobio.
Magnetospirillum (M. magnetotacticum)
Es capaz de responder a campos magnéticos. Es un microorganismo magnetotáctico. Posee partículas de magnetita (entre 5-40), que se disponen unidas en cadena y le sirven para alinearse según las líneas geomagnéticas. En el hemisferio norte se mueven en dirección norte y en el hemisferio sur se mueven en dirección sur. Esta característica puede ser una ventaja evolutiva para mantener estos microorganismos microaerófilos en zonas de baja concentración de oxígeno, cerca de la interfase aerobia-anaerobia (interfase óxica-anóxica).
Azospirillum
Vive en suelos (fundamentalmente en rizosfera). La especie A. lipoferum es de las más estudiadas. Es microaerófila y fija nitrógeno en simbiosis con ciertas plantas tropicales o de climas templados, como cereales. Su asociación con el maíz beneficia a la planta gracias a la fijación del nitrógeno, favoreciendo su crecimiento.
Todos estos organismos forman más de una vuelta completa de la hélice.
Vibrioides
No forman una vuelta completa de hélice. El ejemplo más característico es Bdellovibrio. Es una δ-proteobacteria. El término Bdellovibrio viene del griego “bdella” que significa sanguijuela. Tiene forma de coma, posee un flagelo envainado (vaina=prolongación de la pared celular), lo que facilita su observación, y es de pequeño tamaño.
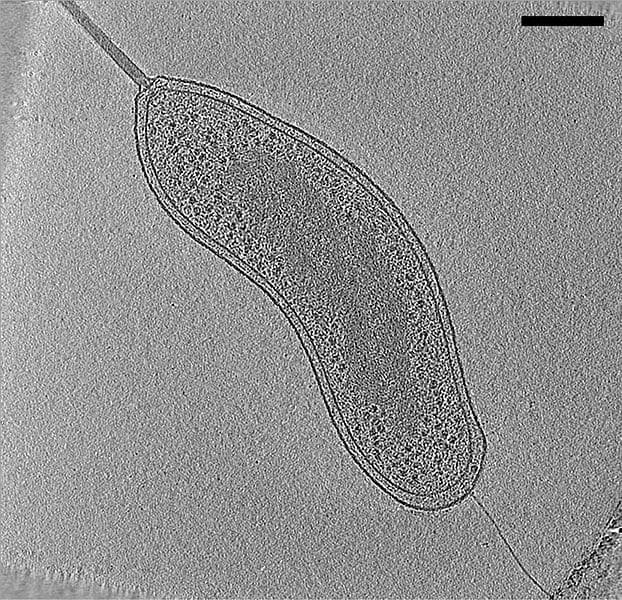
Están presentes en suelos y aguas. Se caracterizan porque depredan a otras bacterias, fundamentalmente GRAM negativas (-).
Bdellovibrio (B. bacteriovorus)
La especie mejor conocida es Bdellovibrio bacteriovorus, la cual tiene un estilo de vida muy característico que alterna dos fases es un ciclo de vida: una fase depredadora extracelular en la que no crece y una fase reproductora intracelular en la que si se divide:
Metabolismo
- Fase extracelular. Es capaz de moverse rápidamente, nada a gran velocidad (10 veces más rápido que otra bacteria móvil), del orden de 70 a 100 veces su longitud por segundo. En principio nada sin dirección preestablecida hasta que resulta atraído por quimiotaxis hasta la presa. Choca con ésta y las dos células unidas giran rápidamente a unas 100 revoluciones por segundo (rps).
- Fase intracelular. Al chocar es capaz de atravesar la pared celular y modificar la membrana externa por la acción de una serie de enzimas hidrolíticos, se desprende del flagelo y taladra la pared celular. Se acomoda en el espacio periplásmico y constituye una estructura esférica más o menos, denominada bdelloplasto. Tarda del orden de 15 a 20 min en acomodarse. Desde esta forma se nutre de todos los componentes de la presa. Inhibe todos los procesos de la otra célula, rompe la membrana citoplasmática y las macromoléculas salen al espacio periplásmico.
Es aerobia estricta y respira aminoácidos y acetato. Es capaz de tomar proteínas enteras, ácidos grasos, aminoácidos y demás. De esta manera economiza tanto carbono como energía, ya que lo toma directamente de la presa. A medida que va biosintetizando se va alargando y finalmente se fragmenta, en tantos fragmentos dependiendo del tamaño de la presa, en E.coli de 5 a 6 Bdellovibrios, en Spirillum de 20 a 30. Una vez fragmentado sintetizan el flagelo y se liberan rompiendo la pared celular de la presa para volver a iniciar el ciclo. Se alejan a gran velocidad. El ciclo puede durar de 2.5 a 4 horas.
Son auténticos depredadores bacterianos. Se supone que Bdellovibrio, junto con protozoos y los fagos, constituyen un modo de control del crecimiento de las poblaciones bacterianas en algunos ambientes.
El modo de ataque y el desarrollo puede clasificarse como único, aunque no es la única bacteria depredadora. Vampirovibrio también se la puede considerar depredadora, en este caso la presa es un alga eucariota, Chlorella, y no pasa al interior del alga, desde fuera la depreda (le ‘chupa’ su contenido). Posee flagelo, pero no envainado.
Detección y aislamiento
La detección y aislamiento de Bdellovibrio se asemeja bastante a la de los fagos. Se hace crecer la presa en placa y se inocula una suspensión de Bdellovibrio filtrada; donde caiga en la placa forma unas calvas en el césped bacteriano. Estos halos generados se diferencian de los halos de los fagos, pues éstos pueden continuar aumentando de tamaño incluso después de que la presa deje de crecer, pudiendo llegar a ocupar casi toda la superficie de la placa. En el caso de los fagos no aumentan de tamaño las calvas. De estos halos se pueden obtener cultivos puros de Bdellovibrio.
No llegan a formar una colonia típica, sino que están en continuo movimiento. Se aprecia una ligera turbidez, pero no colonias en sí. Pude sufrir ataques por fagos, concretamente bdellofagos, virus líticos y de ADN de una cadena.
Proteobacterias con morfología inusual: bacterias con vaina
Son β-proteobacterias y tienen un ciclo de vida especial en el cual hay dos etapas. Pasan de formas envainadas (en zonas ricas en nutrientes) a formas nadadoras sin vaina en busca de nuevos entornos, dejando las vainas abandonadas. Cuando están en forma nadadora desarrollan flagelación lofótrica. Las vainas están constituidas por proteínas y polisacáridos, pero no por peptidoglicano.

En condiciones de crecimiento desfavorable, están en forma de célula nadadora por la presencia de flagelos lofotricos en posición subterminal. Se dispersan en busca de nuevos entornos donde crecer. Cuando las condiciones son favorables, las células pierden el flagelo y crecen vegetativamente dentro de las vainas. Son bacterias aerobias y se encuentran cerca de la interfase óxica-anóxica.
Viven en la interfase en aguas ricas en materia orgánica como cursos de agua contaminados, en plantas de tratamiento de agua residual. Son quimioheterótrofas; al inicio de su estudio se creyó que eran quimiolitótrofas, pero no acoplan reacciones redox de compuestos inorgánicos para obtener energía. Llevan a cabo reacciones catalizadas por proteínas de la vaina que conducen a depósito de óxidos metálicos por encima o por dentro de la vaina en ambientes con hierro o manganeso en forma reducida. El agua anóxica con estos compuestos alcanza a la zona óxica y las vainas pueden acabar recubiertas por hidróxidos u óxidos de hierro o de manganeso, debido a la capacidad de oxidar estos iones. Pero no son quimiolitótrofas por que no acoplan estos procesos de oxidación de compuestos inorgánicos a la obtención de energía.
Sphaerotilus
Vive en aguas muy polucionadas y no puede oxidar manganeso. Junto con Beggiatoa es responsable del engrosamiento en plantas de tratamiento de agua residual ya que incrementan la masa del fango impidiendo que estas aguas sean tratadas correctamente.
Leptothrix
Vive en agua dulce rica en sales minerales, pero no contaminada. Suele vivir con concentraciones altas de hierro soluble, aunque también puede oxidar el manganeso.
Proteobacterias pedunculadas
Muchas son α-proteobacterias, pero no todas. Se denominan así porque todas tienen algún tipo de apéndice o pedúnculo. Se denomina prostecas a las extrusiones de diámetro menos a la célula madura y que contienen citoplasma y está unida por la pared celular. Cuando el pedúnculo no es celular se denomina tallo, como en Galionella. Hay pedunculadas gemantes y no gemantes.
Gemantes
Hyphomicrobium
Se divide por gemación en el extremo de la prosteca, que tiene forma de hifa. Se denomina hifa cuando se trata de una prosteca reproductora. La yema se agranda, forma un flagelo y se libera de la célula madre y se aleja nadando. Después pierde el flagelo, deja de ser nadadora y tras un periodo de maduración forma la prosteca. En ocasiones se pueden formar varias yemas en el extremo de la hifa madre. Puede haber grupos de células conectados por hifas.

Tiene un ciclo de vida con división desigual, con dos tipos de células: una prostecada y otra nadadora.
Es quimioheterótrofa y está muy cerca filogenéticamente de Rhodomicrobium (es una bacteria roja no del azufre). Además, es metilótrofa y desnitrificante. Habita comúnmente en suelos, también presente en agua dulce y marina. Para cultivarla sería preciso un medio con compuestos C1 y nitrato, y puede crecer en medios con muy baja concentración de nutrientes.
No gemantes
Su pedúnculo puede tener función de adherencia, favorecimiento de la absorción de nutrientes (mayor relación sup/vol): es una adaptación evolutiva para los ambientes oligotróficos. Suele vivir en ambientes oligotróficos, como el anterior. También le sirve para reducir la velocidad de sedimentación, ya que al ser aerobios estrictos las prostecas pueden evitar el hundimiento de las células hacia el fondo (zonas anóxicas), donde morirían.
Caulobacter
Es quimioheterótrofa. Se encuentra a menudo en la superficie de ambientes acuáticos con los pedúnculos de varias células hundidas para formar una especie de roseta. En el extremo poseen un botón de anclaje formado por un material adhesivo secretado desde el extremo de la prosteca, lo que permite su unión a otra superficie (sustrato, pared celular, otra bacteria…).
Presenta ciclo vital con división desigual: da lugar a una célula prostecada inmóvil y una nadadora sin prosteca. En este caso, la prostecada se va a dividir y va a dar lugar a células nadadoras flageladas que se alejan de la célula madre. La formación de la prosteca es precursora de la división celular (para que se pueda dividir es necesaria la prosteca). Los tamaños de prosteca varían dependiendo de las condiciones ambientales.

Gallionella
Posee una morfología vibrioide. Es quimiolitoautótrofa. Presenta tallo, es decir, un pedúnculo sin citoplasma. Presenta incrustaciones de hidróxido férrico coloidal que es excretado desde la superficie en forma de filamentos. Obtiene la energía de la oxidación de hierro y manganeso, y fijan CO2 por el ciclo de Calvin. Viven en ambientes con abundante ion ferroso y oxígeno, asociados a las bacterias envainadas, a pH neutro. Viven en la interfase por la inestabilidad del ion ferroso a pH neutro y en presencia de oxígeno.

Mixobacterias deslizantes
Son δ-proteobacterias quimioheterótrofas. Respecto a su ecología, podemos decir que son socialmente activos, interaccionan para formar masas celulares visibles sin la ayuda del microscopio. Además, son terrestres y han sido aisladas del suelo en todo el mundo, desde bosques tropicales hasta la tundra ártica.
Respecto a su morfología, podemos decir que son deslizantes, suelen ser bacilos largos e incluso filamentosos que no presentan flagelos, pero pueden moverse por deslizamiento cuando están en contacto con alguna superficie. Son fructificantes, forman cuerpos fructíferos: estructuras multicelulares complejas que implican comunicación intercelular.
Muestran el ciclo de vida y patrones de comportamiento más complejos de todos los procariotas conocidos. Para ser capaz de codificar esta complejidad, su genoma es muy grande. Myxococcus xanthus posee un genoma que supone 2/3 el tamaño del genoma de una levadura como Saccharomyces.

Ciclo vegetativo
En forma vegetativa son bacilos bastante largos y flexibles que se deslizan sobre distintas superficies dejando un rastro mucoide, de limo. Consiguen los nutrientes lisando otras bacterias. Mientras las condiciones ambientales sean favorables podrán mantener este ciclo, dividiéndose por fisión binaria.
En superficies sólidas se puede observar la formación de colonias con crecimiento radiante, planas con bordes irregulares que se van extendiendo. Visto al microscopio se puede apreciar el rastro mucoide. Avanzan en pequeños grupos de células de forma coordinada, de modo que esto confiere coherencia entre todos los miembros de la colonia, las células pueden seguirles el rastro: unas van avanzando y otras van siguiendo ese rastro y agrupándose. Suelen ser colonias coloreadas por la presencia de pigmentos de tipo carotenoide (carotenoides glucosídicos). La luz estimula la formación de los pigmentos carotenoides y una de sus funciones es la fotoprotección.
Proceso de esporulación
Se inicia cuando se agota su aporte de nutrientes. Las células se van agregando y experimentan un proceso de diferenciación formando finalmente el cuerpo fructífero. Es un proceso de desarrollo complejo desencadenado por la inanición y muy controlado por ciertas señales. Este cuerpo fructífero a veces es pedunculado, se distinguen tallo y cabeza.
La mayoría, en el caso de tener cabeza y tallo, la mayoría de las células se acumulan en la cabeza y se transforman en mixosporas. En cada cuerpo puede haber del orden de 109 mixosporas. La transformación de células vegetativas a esporas se denomina maduración. Son en realidad exosporas contenidas en el cuerpo fructífero, que como tales resisten ciertas condiciones adversas como la desecación, radiación ultravioleta, sonicación, privación de nutrientes. No resisten tan bien el calor como endosporas. La morfología, tamaño de mixospora, color, varía dentro de cada género y de cada especie.
En algunos géneros existen los microcistos, que son mixosporas capsuladas. En otros pueden ser cistos, que son varias mixosporas encerradas en una estructura de pared.
Si las mixosporas entran en contacto con nutrientes, germinan, forman células vegetativas que se deslizan y forman colonias. Son todas quimioheterótrofas aerobias con metabolismo respiratorio. La mayoría puede usar aminoácidos como fuente principal de carbono, nitrógeno, energía y fuente de poder reductor.
Poseen enzimas hidrolíticos que rompen la pared de las otras bacterias para liberar los nutrientes. Absorven los productos solubles de esa lisis, sobre todo péptidos pequeños. Se piensa que la organización en cuerpos fructíferos puede ser ventajosa ya que la masa de células es capaz de producir mayor cantidad de enzimas para digerir a la presa más fácilmente de lo que lo puede hacer la célula individual.
Las células vegetativas pueden convertirse directamente a mixosporas por ciertos inductores químicos, como una elevada concentración de glicina.
Clasificación
Se clasifican por funciones o características morfológicas (de la célula vegetativa, mixospora, cuerpo fructífero). La forma de las células vegetativas distingue dos bloques. Las familias Myxococcaceae, Archangiaceae y Cystobacteraceae poseen extremos fusiformes, los miembros de la familia Polyangiaceae poseen extremos redondeados. Stigmatella posee pedúnculo en el cuerpo fructífero y un tallo ramificado. Nannocystis carece de los carotenoides típicos de las mixobacterias. Es una de las bacterias más frecuentes en el suelo. Produce, como muchos actinomicetos, geosminas. Es celulolítico y agarolítico (degrada celulosa y agar).
Bacterias reductoras del sulfato y azufre
Son δ-proteobacterias. Respecto a su metabolismo, podemos decir que son anaeróbicas estrictas, emplean como donador de electrones (fuente de poder reductor) compuestos orgánicos, pero es muy frecuente que sean productos de la fermentación de otros microorganismos como el lactato. Hay algunas que pueden usar hidrocarbonos e incluso el petróleo crudo como donador de electrones.
Impacto en el medio
Están presentes en ambientes acuáticos y terrestres que se han podido volver anóxicos como resultado de procesos microbianos de descomposición; por ejemplo, se encuentran en sedimentos anóxicos con abundante materia orgánica (fondos de pantanos, lagos, etc). El proceso produce sulfatos reducidos (sulfhídricos o sulfuros), y eso produce una capa de bacterias verdes y rojas de azufre que utilizaran el sulfhídrico producido por las sulfatorreductoras de los fondos de los pantanos. De esta manera se establece un ciclo anaeróbico del azufre.
Producen al año millones de toneladas de sulfhídrico. Tiene un olor desagradable a huevos podridos. Puede producir la precipitación de sulfuro de hierro dando un color negro. En zonas costeras, estas áreas se vuelven inhabitables tanto por el olor como por el efecto tóxico del sulfhídrico. En suelos encharcados puede ocasionar la muerte de animales, plantas y otros microorganismos. Es un peligro potencial porque se puede combinar con el hierro (Fe) de citocromos de cadenas de transporte electrónico de otros seres vivos, así como con otros compuestos celulares esenciales que contienen hierro.
Ejerce un impacto negativo a nivel de industria porque causa considerables pérdidas económicas al provocar o acelerar la corrosión anaeróbica de tuberías de hierro. Con las arqueas metanogénicas también tenemos este problema. También pueden encontrarse en el rumen de rumiantes y tracto intestinal de hombre y otros animales.
Clasificación
Hay dos tipos de bacterias que pueden reducir el sulfato, dependiendo de si oxidan el acetato o no:
- Tipo I: Desulfovibrio tiene forma de bacilo curvado con flagelación polar. Es de tipo I, no es capaz de oxidar el acetato y lo excretan.
- Tipo II: la mayoría son de origen marino. Son capaces de usar el acetato y otros ácidos orgánicos como única fuente de carbono, oxidándolos hasta CO2 y H2. Desulfobacter emplea el ciclo TCA modificado (CAC) para obtener ATP adicional. En estas bacterias es crucial el ciclo del H2 para la formación de un gradiente protónico para lo obtención de ATP. Presenta un flagelo polar único (aunque no todas las especies son móviles). Desulfuromonas no reduce sulfatos pero sí azufre. Posee forma de bacilo recto y flagelación subpolar.


Proteobacterias del grupo epsilon
Campylobacter
Tiene forma de espirilo, microaerófilo con tendencia a ser anaerobio (viene a ppO2 muy baja). Móvil por flagelación monotrica polar. Podemos destacar dos especies:
- Campylobacter fetus. Provoca abortos en animales domésticos (ganado ovino, bovino,…)
- Campylobacter jejuni. Puede causar brotes de enteritis transmitidas por alimentos, en ocasiones se aísla con más frecuencia como causa de diarrea que Salmonella o Shigella.

Helicobacter
Es un espirilo microaerófilo que puede presentar de 1 a 6 flagelos en un polo. La especie más conocida es H. pylori, se asocia con casos de gastritis y úlceras pépticas en humanos. Incluso se ha considerado como causa de cáncer de estómago, ya que se ha visto que existe una correlación positiva entre los casos de cáncer gástrico e infección por H. pylori. Este descubrimiento supuso el premio nobel en 2005 a Barry Marshall y Robin Warren.

Está muy extendida entre toda la población mundial. En países en vías de desarrollo aproximadamente del 70-90% de población afectada, en desarrollados del orden de 25 a 50%. Se han hecho estudios que relacionan determinadas cepas con determinadas migraciones humanas a nivel mundial.
El modo exacto de transmisión no está del todo claro, aunque se especula que puede ir de individuo a individuo, fundamentalmente durante la infancia, aunque hay quien sostiene que es a través de agua y alimentos.
Coloniza estrictamente las células gástricas secretoras de mucus, por debajo de la capa del mucus gástrico. En este lugar produce inflamación que puede llevar consigo una úlcera. Se adhiere por medio de fimbrias. Entre los factores de virulencia podemos encontrar grandes cantidades de ureasa producidos por esta bacteria, la cual degrada la urea a CO2 + amoníaco. La hidrólisis de la urea está asociada con su virulencia. El amoníaco basifica el medio y podría proteger al microorganismo del pH ácido del estómago. Sin tratamiento antibiótico adecuado puede llegar a destruir el tejido y producir una úlcera.

Bibliografía de imágenes
Bouchal, T., Závada, J., Vojtková, H., Langarová, S., & Havelek, R. (2012). Primary Research of Mine Waters from the Chrustenice Iron-Ore Deposit/ Prvotní Průzkum Důlních Vod Železorudného Ložiska Chrustenice. GeoScience Engineering, 58, 23 – 27. DOI:10.2478/gse-2014-0041
Fung C, Tan S, Nakajima M, Skoog EC, Camarillo-Guerrero LF, Klein JA, et al. (2019) High-resolution mapping reveals that microniches in the gastric glands control Helicobacter pylori colonization of the stomach. PLoS Biol 17(5): e3000231. https://doi.org/10.1371/journal.pbio.3000231
Hylemon, Phillip. (1973). Designation of Spirillum volutans Ehrenberg 1832 as Type Species of the Genus Spirillum Ehrenberg 1832 and Designation of the Neotype Strain of S. volutans: Request for an Opinion. International Journal of Systematic and Evolutionary Microbiology. 23. https://doi.org/10.1099/00207713-23-1-20
Kuever, Jan; Rainey, Fred A.; Widdel, Friedrich. (2015). Desulfuromonas. Bergey’s Manual of Systematics of Archaea and Bacteria. (John Wiley & Sons, Ltd): 1-7. https://doi.org/10.1002/9781118960608.gbm01011
Pfennig, N., & Biebl, H. (1976). Desulfuromonas acetoxidans gen. nov. and sp. nov., a new anaerobic, sulfur-reducing, acetate-oxidizing bacterium. Archives of microbiology, 110(1), 3–12. https://doi.org/10.1007/BF00416962
PLoS Biology Issue Image | Vol. 3(10) October 2005. PLoS Biol 3(10): ev03.i10. https://doi.org/10.1371/image.pbio.v03.i10
PLoS Biology Antisocial Behavior in Cooperative Bacteria (or, Why Can’t Bacteria Just Get Along?). November 2005. PLoS Biol 3(11): e398. https://doi.org/10.1371/journal.pbio.0030398


