
La célula procariota
arqueas, bacterias, célula, procariotas | 31 de marzo de 2021 | 36 min de lectura | ¿Prefieres oír el artículo? ¡Dale al ▶︎!
Cargando...
Configuración
Las células procariotas, como las bacterias, son organismos unicelulares caracterizados por su estructura sencilla y su tamaño variable. Este tamaño influye en su eficiencia metabólica y en la velocidad de crecimiento, lo que les permite adaptarse a distintos entornos.
Además, su morfología diversa, desde esféricos cocos hasta alargados bacilos y espirales espirilos, juega un papel importante en su clasificación taxonómica.
Índice de contenidos
Tamaño de la célula procariota
Para calcular el tamaño de un organismo procariota se utiliza un microscopio especial con una regla calibrada. Es característico para cada tipo de microorganismo; además, este factor tiene valor taxonómico. Por ejemplo, Escherichia coli mide entre 0,5-1 micras x 3-5 micras (anchura x longitud). Hay células bacterianas con menos de una micra (como Bdellovibrio con 0,1 micras) o que miden mucho más (Oscillatoria, con hasta 50 micras, o Epulopiscium, con hasta 600 micras). En las células esféricas se mide el diámetro.
El tamaño es importante para estos factores:
- La relación superficie de la célula con respecto al volumen interior. Si es muy alta favorece que el intercambio de nutrientes de la célula con el medio sea muy eficaz (las células son muy eficaces en el transporte y consumo de energía). La velocidad de capturar alimentos es inversamente proporcional al tamaño celular; mejor cuanto más pequeño sea el organismo.
- La velocidad o tasa de crecimiento. Cuanto más pequeña sea una célula, contendrá menos estructuras internas, y su tasa de división será más rápida. Oscillatoria y Epulopiscium, al tener mayor tamaño, tardan más en dividirse.
Forma, morfología de la célula procariota
Se encuentran 3 patrones básicos de morfología en los organismos procariotas:
- Células esféricas o cocos
- Células alargadas o bacilos
- Células en espiral o espirilos
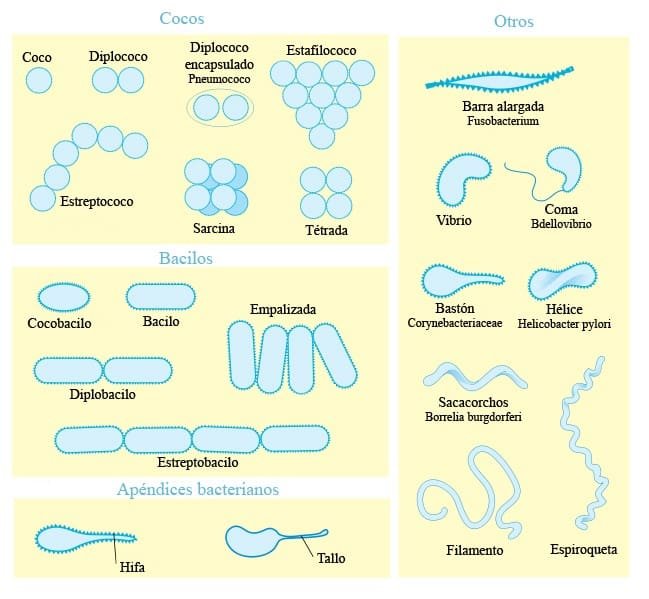
Ahora bien, pueden aparecen más formas. Si encontramos una repetición continua de espirilos, hablamos de espiroquetas. También hay bacterias que presentan apéndices muy largos; estas serían las bacterias pedunculadas (apéndice = pedúnculo). Además, hay algunas bacterias con disposición miceliar; son los actinomicetos o actinobacterias.
En el caso de las células con morfología de coco, cuando una célula se divide en un solo plano la célula hija queda unida a la madre, sin separarse. Si esto se repite n veces, nos queda un conjunto de células unidas formando una cadena, formando la morfología de ‘estrepto’ y que define al género Streptococcus. Si en lugar de en un plano, el coco se divide en planos múltiples con un patrón irregular, obtenemos una masa de células formando un conjunto que recuerda a un racimo de uvas, formando la morfología de ‘estafilo’ y que define al genero Stafilococcus. En el caso de Corynebacterium dyphteriae (produce la difteria) podemos identificarla porque forma bacilos en empalizada muy característicos del genero Corynebacterium.
Envoltura procariota
Vamos a ver las capas de la envoltura procariota desde un orden de dentro a fuera.
Membrana plasmática
La membrana plasmática es una capa fina que rodea a la célula. Es una barrera osmótica esencial que regula el tráfico de nutrientes y productos entre la célula y el medio. Mantiene la misma estructura básica de todas las membranas biológicas. Su estructura básica está formada por una bicapa lipídica con disposición de fluido móvil donde las cabezas polares están hacia el exterior de la membrana y las colas hacia el interior. Químicamente, no aparecen los mismos fosfolípidos que en eucariotas, pero la disposición es la misma. Hay dos propiedades a destacar:
- Contienen una gran cantidad de actividades enzimáticas. Por ejemplo, cuentan con permeasas (transportan y controlan el paso de sustancias), enzimas biosintéticas (sintetizan los componentes celulares propios), ATPasas (cadenas de transporte de electrones), enzimas que intervienen en procesos de señalización con el medio y, en muchas bacterias fotosintéticas, parte del complejo fotosintético se encuentra en la membrana. Las membranas de procariotas son metabólicamente mucho más activas que las de eucariotas, ya que no hay orgánulos y la membrana realiza el trabajo de estos orgánulos ausentes. Las proteínas de membrana de procariotas son generalmente transmembranales, y tienen dominios hidrofóbicos para quedar anclados a los fosfolípidos, además de dominios tanto dentro como fuera de la célula.
- No tienen esteroles. Esto los diferencia de las células eucariotas, donde si aparecen esteroles. En su lugar tienen ergosteroles, o en el caso de algunos procariotas, hopanoides que se parecen al colesterol.
Mesosomas
Los mesosomas son invaginaciones o repliegues que se producen junto al tabique de división celular cuando la célula va a dividirse por fisión binaria. Son puntos de anclaje para el nucleoide bacteriano de la célula en división. Son más numerosos en GRAM+ que en GRAM-. Hay algunas especies, como Azotobacter, con una elevada actividad respiratoria (la tasa de respiración celular más alta de todos los seres vivos), que tienen una gran cantidad de mesosomas.
Pared celular
La pared celular es la capa externa que rodea a la membrana. Le produce la forma característica de dicho procariota, le da protección osmótica, y le confiere una elevada rigidez. En la mayor parte de procariotas es la capa más externa, si bien no en todas es así. La pared celular es una estructura esencial; de hecho, si la bacteria la pierde, se lisa y se muere. Todos los organismos procariotas tienen pared a excepción de los micoplasmas. A destacar:
- La pared celular de bacterias y arqueas es diferente. La pared bacteriana destaca porque está formada por polímeros exclusivos que no se encuentran en ningún otro ser vivo, el peptidoglicano (o mureína).
- La pared es portadora de importantes factores relevantes en la patología bacteriana que producen enfermedades. Además, protege a la bacteria de sustancias toxicas del medio (como los antibióticos) y contra la fagocitosis del sistema inmune y el complemento. La pared bacteriana es la diana sobre la que actúan algunos de los antibióticos más efectivos, como la penicilina.
- Las diferencias en la composición química y en la estructura de la pared celular permiten clasificar a los organismos procariotas en dos grandes grupos: GRAM+ y GRAM-.
Peptidoglicano
Este es un resumen sobre el peptidoglicano. Para un análisis más alargado sobre el peptidoglicano (composición, biosíntesis, antibióticos que les afecta) revisa este artículo.
El peptidoglicano es un compuesto de procariotas, exclusivo de bacterias, formado por azúcares y aminoácidos. Es el compuesto cuya función es dar rigidez, consistencia y forma a la pared celular bacteriana. El peptidoglicano está formado por una secuencia alternante de dos azúcares, N-acetil-glucosamina (NAG) y el ácido N-acetilmurámico (NAM), unidos por enlaces β(1→4), entre el C1 de NAM y el C4 de NAG. NAM es un derivado de NAG al que se le ha añadido una molécula de ácido láctico en el C3. La estructura del peptidoglicano está conformada por tres capas que veremos más adelante. Hay que destacar que, tanto en hongos como en las cubiertas de crustáceos o moluscos, también hay N-acetilglucosamina.
La primera capa del peptidoglicano (horizontal) está formada por filas lineales de NAG y NAM unidos de forma secuencial, mediante enlaces covalentes.
La segunda capa (vertical) está formada por un tetrapéptido unido solamente a NAM (esta es otra diferencia entre NAG y NAM). A este tetrapéptido se le denomina tetrapéptido del glicano. Este tetrapéptido contiene:
- L-alanina
- D-glutámico (en algunas bacterias puede estar hidroxilado)
- L-lisina en la mayoría de GRAM+ o meso-diamino-pimélico (m-DAP) en GRAM- y algunas GRAM+ (en GRAM+ también pueden aparecer otros aminoácidos)
- D-alanina
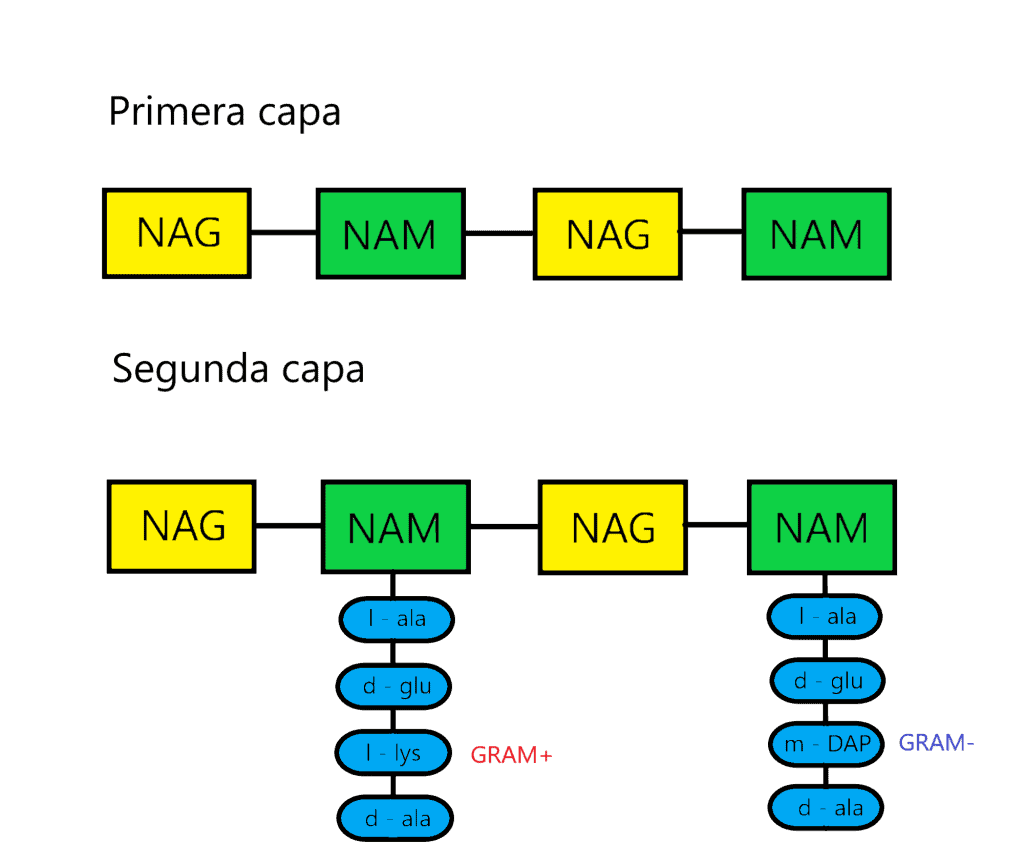
Como se puede ver, aquí aparecen proteínas formadas tanto de d-aminoácidos como de l-aminoácidos, de ahí una de sus particularidades (ya que las células normalmente usan solo l-aminoácidos para formar proteínas).
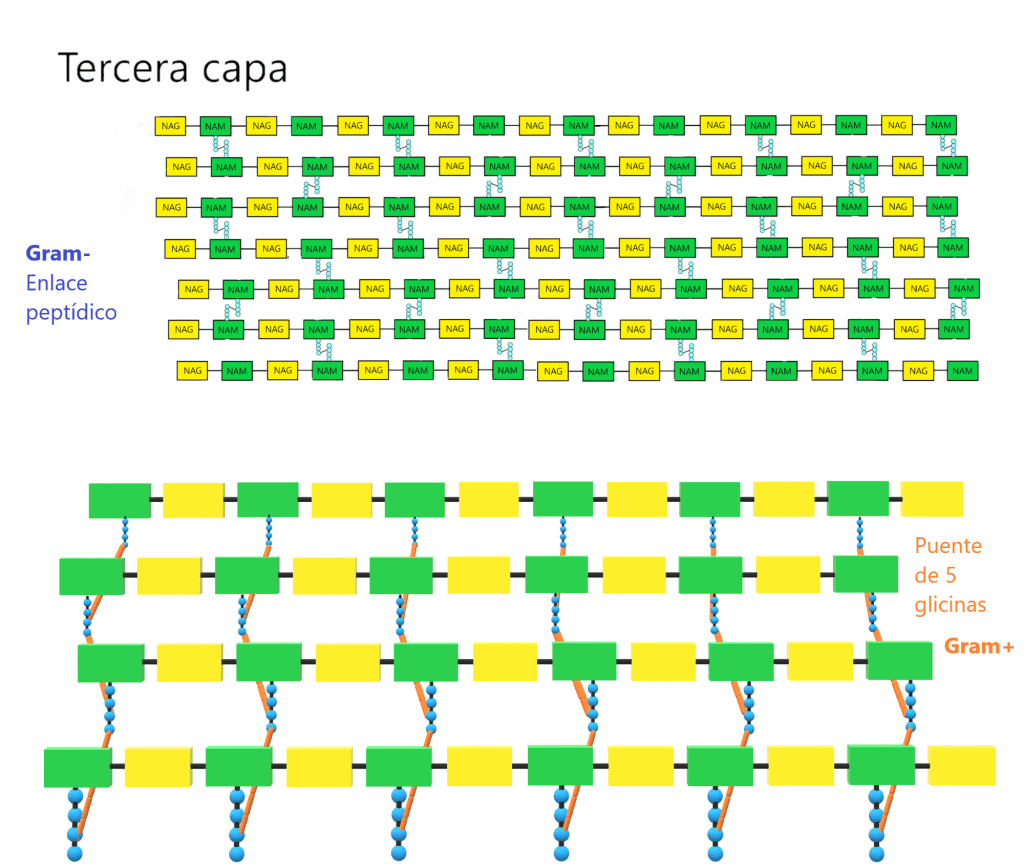
La tercera capa está formada por la unión de dos NAM vecinos. Para ello, la mayoria de veces se forma un enlace peptídico entre el cuarto aminoácido de un tetrapéptido (d-alanina) y el tercero de otro (L-lisina o m-DAP) mediante una reacción de transpeptidación. La composición de estos puentes peptídicos varía entre los diferentes grupos bacterianos; de hecho, los aminoácidos y la posición de este enlace se ha usado para clasificarlos (taxonomía). En bacterias GRAM- y algunas especies de Bacillus el enlace es directo, entre la D-alanina y el ácido diaminopimélico (m-DAP). En la mayoría de bacterias GRAM+ hay un puente formado por uno o más aminoácidos. La penicilina ataca esta etapa, la transpeptidación, inhibiendo su formación.
Estas tres capas forman la estructura tridimensional que da rigidez a la pared. Aunque pueden haber variaciones en la parte proteica del peptidoglicano, los enlaces entre NAG y NAM iguales para todas las especies bacterianas. No todos los tetrapéptidos unidos a NAM sufren la reacción de transpeptidación. De hecho, la pared tiene que tener algo de porosidad para permitir el paso de sustancias por la pared. Además, para que la célula pueda replicarse, necesita dividirse, expandirise, y es otra razón por la que la pared debe tener cierta flexibilidad. Ahora bien, cuantos más tetrapéptidos sufren la reacción de transpeptidación, más resistente será la pared celular.
Biosíntesis del peptidoglicano
La síntesis del nuevo peptidoglicano ocurre durante el proceso de crecimiento celular. Hay cuatro fases en este proceso. Primero, se forman los precursores del PG, que son derivados UDP de aminoazúcares formados en el citosol. Después, los aminoazúcares se transfieren a un transportador lipídico de membrana (bactoprenol) que los lleva a los aminoazúcares a través de la membrana celular. Más adelante, el PG se formará en la superficie externa de la membrana. Por último, ocurrirá la reacción de transpeptidación que forma puentes entre tetrapéptidos de NAM.

Para un análisis más exhaustivo sobre el peptidoglicano (composición, biosíntesis, antibióticos que les afecta) revisa este artículo.
Espacio periplásmico
El espacio periplásmico se encuentra entre el peptidoglicano y la membrana (ya sea externa o plasmática). Es una zona metabólicamente muy importante. Muchas de las actividades enzimáticas se localizan aquí. Además permite interconexiones temporales entre las capas.
Diferencias de la pared entre GRAM positivas y GRAM negativas
| Diferencias entre bacterias GRAM+ y GRAM- | ||
|---|---|---|
| Bacterias | GRAM+ | GRAM- |
| Tamaño de PG | Hasta 90% de la pared celular | Hasta el 10% de la pared celular |
| Localización del PG | Al exterior de la membrana plasmática | Entre la mebrana interna y la membrana externa |
| Tinción de GRAM | Complejo cristal violeta-yodo | Safranina |
| Color en la tinción de GRAM | Azul, violeta | Rojo, rosa, naranja |
| Tercer aminoácido del tetrapéptido | L-lisina normalmente; mucha variación de aminoácidos | m-DAP casi siempre; poca variación de aminoácidos |
| Transpeptidación (cross-linking) | Casi siempre puente de aminoácidos | Casi siempre enlace directo |
| Compuestos unidos a PG | Ácidos teicoicos | Lipopolisacárido (LPS) |
En el cuadro de arriba tienes las diferencias entre GRAM+ y GRAM- de forma más resumida. Las mayores diferencias están en la estructura y organización de la pared. Por ello, se tratan con antibióticos diferentes.
GRAM positivas
En bacterias GRAM+, el peptidoglicano es una capa muy gruesa, y ocupa entre el 70-90% de la pared celular. El nivel de transpeptidación es muy alto. Pueden aparecer bacterias con solo una capa de peptidoglicano rodeando a la célula, pero la mayoría de bacterias cuenta con muchas láminas apiladas unas sobre otras. Eso sí, la capa se sitúa en el exterior de la membrana plasmática. En la tinción de Gram, dentro de las células se forma un complejo cristal violeta-yodo que NO puede extraerse. Y es debido a sus gruesa pared celular formada por varias capas de peptidoglicano. Al introducir alcohol, se deshidratan, lo que provoca el cierre de los poros de las paredes, así que el colorante no sale de la bacteria. Además, en bacterias GRAM+ aparecen los llamados ácidos teicoicos. Son polímeros de la pared formados por un azúcar alcohol, que pueden ser unidades de glicerol de 3 carbonos (glicerolfosfato) o de ribitol de 5 carbonos (ribitolfosfato), y unen entre si por grupos fosfato, polímeros largos (glicerolfosfato, ribitolfosfato). También pueden tener unidos otros azúcares y d-alanina. Los ácidos teicoicos se unen mediante enlace covalente a residuos de NAM. Pueden intervenir uniendo calcio (Ca2+) y magnesio (Mg2+) para su transporte. Los ácidos teicoicos tienes varias funciones:
- Compactación y estabilización de la pared
- Fijación del peptidoglicano a la membrana plasmática
- Tienen carga negativa, así que tienen su parte de responsabilidad de la carga negativa de la superficie celular
- Son considerados los principales antígenos superficiales de las GRAM+
GRAM negativas
En bacterias GRAM-, el peptidoglicano ocupa hasta el 10% de la pared celular. No aparecen ácidos teicoicos. Está situada entre la membrana externa y la membrana interna de la célula. La mayor parte de la pared celular está formada por la membrana externa, una segunda bicapa lipídica que, aparte de fosfolípidos y proteínas, también contiene polisacáridos. A esta membrana se le llama lipopolisacárido (LPS).
En la tinción de Gram, dentro de las células se forma un complejo cristal violeta-yodo que puede extraerse. Y es debido a que la membrana externa es rica en lípidos, además de que la capa de peptidoglicano es tan fina que no impide el paso del solvente, así que el colorante sale de la célula. Después del tratamiento con alcohol, estas bacterias reciben una tinción de contraste, la safranina, para diferenciarse de las bacterias GRAM+.
Las GRAM- tienen ciertos componenetes exclusivos tales como:
- Lipoproteína de Brown. Proteína pequeña que encarga de engarzar la membrana externa a la capa de peptidoglicano.
- Uniones físicas espaciales. Se encargan de la interacción entre la membrana externa y la membrana plasmática.
- Lipopolisacárido (LPS). Son endotoxinas de bacterias GRAM-, que producen los cuadros de fiebre. El LPS tiene una función estabilizadora sobre la membrana externa. Además, sus grupos fosfato otorgan carga negativa a la célula.
Hay una enorme variedad química dentro de los organismos GRAM-, pero todos tienen un mismo patrón general que consta de 3 partes: lípido A, al que se une la Región Core, que se continua con una cadena lateral O o tejido somático.
- Lípido A. Es la parte hidrófoba. Su función en la de anclar el conjunto del lipopolisacárido a la membrana externa. Está formado por dos residuos de glucosamina fosforilados a los que se anclan 2 o 3 cadenas (ramificadas o no) de ácidos grasos.
- Región Core. Es una parte glucídica en la que aparecen, junto a azucares comunes, dos azucares especiales, heptosas, que son la heptulosa y el KDO (ácido 2-keto-3-deoxioctanato). Se unen a través de un enlace fosfato a los fosfolípidos de la membrana. Son importantes porque a ellos se une la etanolamina a un residuo fosforilado.
- Cadena lateral o antígeno «O». En esta cadena aparecen azucares inusuales como la ramnosa, la tivelosa o la abecuosa. Es muy común que los azucares se dispongan en pentasacáridos o tetrasacáridos, que se repiten n veces. Esta cadena lateral se denomina antígeno O, ya que genera una reacción de anticuerpos o linfocitos T (reacción inmune). Al incluir los pentámeros, se modifica la composición de azucares de la cadena lateral, lo que provoca que los anticuerpos no los puedan reconocer.
Las GRAM-, en general, son mucho más resistentes que las GRAM+ al ataque de los antibióticos, los desinfectantes y a la acción de otras moléculas toxicas. Esto ocurre debido a que la membrana externa es más hidrofóbica, y por lo tanto, impermeable al paso de sustancias. Las GRAM- necesitan una permeabilidad selectiva para el paso de nutrientes, y se lleva a cabo por porinas (3 unidades que se insertan en la cadena externa y forman un poro o canal, por la cual pasan moléculas de 600-700 Dalton). Para tamaños mayores hay otros sistemas de transporte.
Cápsulas bacterianas
La cápsula es una capa de naturaleza viscosa o mucilaginosa que presentan algunos procariotas externamente a la pared celular. No es indispensable para la vida del organismo procariota, pero si que le es útil para resistir ataques del sistema inmune o de antibióticos. El glicocálix es una red de exopolímeros (polisacáridos o polipéptidos) que rodean a la célula bacteriana. Se denomina cápsula cuando el material externo está organizado y firmemente unido a la pared celular. Se denomina Slime o capa mucilaginosa cuando el material es difuso y no organizado. Hay bacterias que pueden perder la cápsula o no formarla, y ha bacterias que dependiendo del medio donde crezcan la naturaleza de su cápsula será distinta. Al no ser una estructura esencial, la bacteria puede crecer, dividirse y perder la cápsula sin verse perjudicada.
Estas son las funciones de las cápsulas:
- Resistencia a la desecación, ya que previene la deshidratación por su capacidad de retención de agua.
- Evita la fijación de fagos y materiales tóxicos hidrofóbicos.
- Es una fuente de nutrientes cuando las condiciones del medio son desfavorables.
- Puede ayudar en la movilidad. Así, aparece el slime en bacterias deslizantes.
- Es un factor de virulencia en algunas especies, y las protege de la fagocitosis o de los antibióticos.
- Mediador de interacciones celulares.
Tipos de cápsulas
Según su composición, la cápsula puede ser:
- Polipeptídica. Algunas bacterias del genero Bacillus (Bacillus anthracis) sintetizan una capsula externa formada por unidades de D-glutámico.
- Polisacárida. Son las capsulas más comunes. Hay dos tipos:
- Con polisacáridos de origen interno. Constituidas por polisacáridos sintetizados directamente por la célula. Se sintetizan dentro y se secretan a la pared. Hay gasto de ATP para formar la cápsula, utilizando precursores de su propio metabolismo para su formación. Estas capsulas están muy extendidas. No dependen de las condiciones del medio. El polímero puede ser de:
- Celulosa. Acetobacter xylinum.
- Glucano. Agrobacterium tumefaciens.
- Ácido colánico. Enterobacterias.
- Poliurónidos. Pseudomonas aeruginosa.
- Polisacáridos pneumocócicos (varios tipos). Streptococcus pneumoniae.
- Con polisacáridos de origen externo. Se forma a partir de la sacarosa o azúcar de caña (uno de los azúcares más abundantes). No se gastan ni sus precursores ni su ATP, pero dependen de las condiciones del medio. Hay dos tipos:
- Levanos. Se añade a una molécula de sacarosa una fructosa vecina, liberándose una glucosa. Ocurre en algunas especies de Bacillus, Pseudomonas y Xhantomonas
- Dextranos. Se añade a una molécula de sacarosa una glucosa, liberándose una fructosa. Ocurre en Leuconostoe y Streptococcus mutans (bacteria que provoca caries).
- Con polisacáridos de origen interno. Constituidas por polisacáridos sintetizados directamente por la célula. Se sintetizan dentro y se secretan a la pared. Hay gasto de ATP para formar la cápsula, utilizando precursores de su propio metabolismo para su formación. Estas capsulas están muy extendidas. No dependen de las condiciones del medio. El polímero puede ser de:
Características de la membrana y pared de las arqueas
En la membrana hay ciertas diferencias respecto a los fosfolípidos constitutivos. Aparece el modelo de mosaico fluido y bicapa lipídica. Los fosfolípidos son del grupo de los hidrocarburos alifáticos, del grupo de los fitanoles. A diferencia de las bacterias, en arqueas los fosfolípidos se unen al glicerol por un enlace tipo éter, que es mucho más rígido. Aunque no aparece siempre, es relativamente común en arqueas que, en lugar de una bicapa lipídica, aparezca una única monocapa.
La diferencia más destacada es la ausencia de peptidoglicano en su pared. Eso si, poseen una gran variedad de posibilidades: desde sin pared hasta las que tienen la llamada capa S. Algunas tienen una sustancia parecida al peptidoglicano que se denomina pseudopeptidoglicano, formado por N-acetil-glucosamina unida a N-acetiltalosaminomurámico mediante enlaces β(1→3), inatacable por la lisozima.
- Las llamadas capas S aparecen tanto en arqueas como en bacterias y están constituidas por proteínas ricas en aminoácidos azufrados (cisteína y metionina) y adoptan un patrón en mosaico. En GRAM+ aparecen adheridas al peptidoglicano. En GRAM- adheridas a la membrana externa. En el caso de las arqueas, las capas S pueden ser la única estructura tipo pared que rodea a la membrana plasmática, de ahí que su importancia sea mayor en arqueas que en bacterias.
Las funciones de las capas S son:
- Contribuir a la forma y a la rigidez de la célula.
- Parece ser que también intervienen en la protección frente a cambios ambientales de pH o de contenido de agua.
- Aumentan la virulencia
- Mejoran la adhesión celular a la superficie.
Flagelos
La gran mayoría de seres vivos unicelulares viven en ambientes que no son del todo líquidos, sino con un cierto grado de viscosidad. Necesitan desplazarse en estos ambientes, y para ello presentan una estructura denominada flagelo (en eucariotas hay una estructura equivalente con otro nombre, undulipodio).
Los flagelos son apéndices de tipo filamentoso, muy largos y finos, semirrígidos. Se encuentran por debajo del límite de resolución de los microscopios convencionales. Hay algunas tinciones que incorporan acido tánico para verlos al microscopio, aunque es muy problemático porque al fijar las preparaciones el flagelo se pierde. Son estructuras que están ancladas a la membrana plasmática de la célula y desde allí, atravesando la pared, se extienden hasta el medio externo. El filamento puede llegar a medir hasta 10 veces el diámetro celular y está constituido por una proteína exclusiva llamada flagelina. Hay diferencias o variaciones entre distintas especies, pero todas las flagelinas son ricas en aminoácidos diácidos (aspártico y glutámico).
Los flagelos bacterianos tienen carácter antigénico; de hecho, el antígeno H es flagelar. Los flagelos más gruesos se deben a la presencia de una envoltura membranosa que rodea al flagelo. Este tipo de envoltura es característico de cada especie y algunas bacterias, dependiendo de la naturaleza del medio, pueden formar o no envoltura.
Estructura de un flagelo
Desde fuera hacia dentro está formado por estas 3 partes:
- Filamento helicoidal. Se proyecta hacia el exterior y es semirrígido. Está constituido por subunidades de flagelina que tienen la capacidad de autoensamblarse. La flagelina es una proteína que se sintetiza por los ribosomas; a medida que se sintetizan, viajan por el hueco interno del flagelo, y cuando llega al extremo apical hay un cambio conformacional en la flagelina que provoca que las unidades se depositen como si fueran ladrillos. Hay gasto de energía en la síntesis de flagelina, pero no es su ensamblaje.
- Gancho. Presenta una amplitud ligeramente superior. Está más próximo a la superficie de la célula. Sirve de mango de unión entre el filamento y el cuerpo basal. Su función es transmitir el movimiento.
- Cuerpo basal o rotor flagelar. Está dentro de la pared, anclado a la membrana plasmática. Consta de un conjunto de proteínas necesarias para el movimiento; de hecho, es la zona donde ser genera el movimiento del flagelo. Consta de un sistema complejo de anillos cuya estructura varía dependiendo de si la bacteria es GRAM+ o GRAM-.
- En GRAM- hay dos pares de anillos. El par más externo se denomina par L-P: L (insertado en la zona correspondiente al LPS) y P (insertado en el peptidoglicano). Este par tiene función estructural y de fijación. El par de anillos mas interno es el par S-M: S (en el espacio periplásmico) y M (en la membrana). Debajo del anillo M hay un quinto anillo, el anillo C, que está en contacto con el par S-M y se encarga de regular el giro del flagelo.
- En GRAM+ el par externo no existe. Solo encontramos el interno, el par S-M, y el anillo C. El par S-M es el encargado del movimiento. El L-P es solo estructural.
Clasificación de los flagelos
Atendiendo al número de flagelos y a su disposición, la flagelación puede ser:
- Flagelación polar. Un solo flagelo en uno de los extremos de la célula, como Pseudomonas o Vibrio. También se denomina flagelación monotrica o subpolar.
- Flagelación lofotrica. Un grupo de flagelos dispuestos en uno de los polos de la célula, como Spirillum.
- Flagelación anfitrica. Tienen un solo flagelo en cada uno de los dos extremos opuestos. En este caso, un solo flagelo opera a la vez, lo que permite a la bacteria revertir su movimiento de forma rápida activando el flagelo opuesto.
- Flagelación peritrica. Todos los flagelos se disponen rodeando el perímetro exterior de la célula, como algunas las cepas móviles de E. coli.
Bases del movimiento flagelar
Los flagelos actúan como un rotor de hélice, un rotor helicoidal semirrígido. No pueden flexionarse. En conjunto, todos los flagelos se mueven girando como una hélice sobre su eje. El movimiento flagelar se genera en la base, donde está el cuerpo basal, y desde ahí se transmite hacia el filamento, hasta el gancho. Para que el motor flagelar gire debemos contar con el par S-M operativo. Ambos anillos rotan; uno gira sobre el otro. El proceso de movimiento requiere energía que procede de la fuerza protón-motriz que se genera en la membrana cuando se disipa el gradiente de la membrana. El movimiento es un movimiento muy controlado siempre. En cada momento, la bacteria decide la velocidad de rotación, la orientación, y decide la frecuencia de movimiento y parada. Generalmente se produce a favor o en contra de gradiente. Esto tiene que ver con el flagelo típico bacteriano, que es externo.
El movimiento de la bacteria a través del flagelo se denomina taxis. La taxis siempre es a favor de gradiente. Cuando se mueve a favor de gradiente de sustancias químicas se denomina quimiotaxis. Si la bacteria se mueve a favor de un gradiente de luz se denomina fototaxis. Si lo hace a favor de un gradiente de oxígeno se denominan aerotaxis.
Hay un grupo específico de bacterias, las espiroquetas, que tienen un filamento axial. Son endoflagelos. Parten de los extremos de la célula y se proyectan hacia el interior. Lo que hacen es facilitar un movimiento de tipo reptante, mediante contracción-extensión.
Pelos y Fimbrias
En bacterias tenemos otros tipos de apéndices más cortos denominados pelos y fimbrias. Ambos términos se utilizan de forma indistinta, pero no son lo mismo. Eso si, ninguno de los dos interviene en el movimiento.
- Los pelos son un poco más largos y aparecen en menor proporción que las fimbrias. Tienen una función sexual: la conjugación bacteriana. Están formados por una proteína exclusiva denominada pilina.
- Las fimbrias son mucho más cortas y más abundantes. No son huecas. Las bacterias de vida libre poseen numerosas fimbrias, pero al cultivarlas en el laboratorio las pierden. Su función en la de reconocimiento del sustrato y adherencia. Permiten que las bacterias colonicen hábitats muy específicos, como la mucosa intestinal. Este proceso de adherencia es importante para las bacterias que van a producir procesos invasivos.
Organización interna de la célula procariota
El citoplasma de procariotas, al no tener orgánulos, se ve como una especie de matriz más o menos homogénea, de aspecto granular debido a la abundancia de ribosomas libres. En los ribosomas, el proceso de transcripción y traducción está acoplado: los ribosomas se unen al mRNA (ARN mensajero) y lo van leyendo, a la vez que sintetizan proteínas. Es muy común observar, durante la fase activa de síntesis de proteínas, a los ribosomas como un grupo de partículas enlazadas a un mRNA en crecimiento formando los polirribosomas. En procariotas, el nucleoide o genóforo es una doble hélice de ADN sin aislamiento de ninguna membrana y carente de proteínas básicas que lo estabilicen como las histonas.
Aunque no tengan orgánulos, si que tienen estructuras especiales, como las que veremos a continuación.
Inclusiones de reserva
Aparecen dentro del citoplasma como cuerpos de inclusión. Son gránulos que pueden teñirse y verse al microscopio (de ahí su nombre de inclusión). Los procariotas son capaces de acumular reservas orgánicas e inorgánicas, pero no acumulan grasas neutras. Hay diversos tipos:
Inclusiones de reserva de carbono
Son las más típicas. Las más importantes son los polímeros de glucosa denominados glucanos. Dentro de los glucanos encontramos:
- Glucógeno. Compartido por procariotas y muchos eucariotas. Formado por unidades lineales de glucosa unidas por un enlace α(1→4) y con ramificaciones en α(1→6)). El glucógeno aparece como gránulos internos.
- Almidón. En plantas. Hay muy pocos procariotas que acumulen almidón.
El otro polímero que sirve como reserva de carbono (exclusivo de procariotas, tanto bacterias como arqueas) es el PβHB (Poli-β-Hidroxibutirato), elemento que nunca se ha encontrado en eucariotas. Está formado por unidades repetitivas del ácido β-Hidroxibutírico unidos mediante un enlace éster entre el grupo carboxilo de un ácido y el hidroxilo del siguiente. Tiene carácter neutro y para visualizarlo se emplea un colorante (tinción de negro sudán).
Normalmente cada especie bacteriana acumula una única inclusión de reserva. Por ejemplo, las enterobacterias (como E. coli) y el género Clostridium acumulan glucógeno. En cambio, Pseudomonas y Bacillus acumulan ácido polihidroxibutírico (PHB). Dentro de las Pseudomonas, Pseudomonas florescens no acumula ninguna sustancia de reserva.
Inclusiones de reserva de nitrógeno
En general, los procariotas no acumulan reservas de nitrógeno orgánico, a excepción de la cianoficina, que es sintetizada por las cianobacterias. En ellas, el exceso de nitrógeno que no consumen lo acumulan en forma de cianoficina, un polímero formado por dos aminoácidos en igual proporción: aspártico y arginina. La cianoficina se sintetiza cuando la bacteria se encuentra en fase estacionaria (cuando no está creciendo); si luego necesita activar su metabolismo otra vez se movilizan estas inclusiones.
Cuando se bloquea la síntesis de proteínas en los ribosomas, estas cianobacterias pueden seguir sintetizando cianoficina. Esto indica que no se sintetiza en los ribosomas.
Gránulos de polifosfato o volutina
Se acumulan en forma de fosfato inorgánico. Se les denomina corpúsculos metacromáticos: esta sustancia teñida sufre un proceso de inversión cromática. Se sintetizan cuando hay alguna limitación de un nutriente, sobre todo de sulfatos. Se sintetizan a partir de ATP, mediante la transferencia de un fosfato de alta energía a un polímero de volutina.
ATP + P(n) → P(n+1) + ADP
Estas acumulaciones tienen como objetivos:
- La célula acumula fosfato para movilizarlo cuando tenga que replicar los ácidos nucleicos.
- Fuente de energía. Esto supone que la reacción tiene que ser reversible, para volver a formar ATP a partir de las reservas de fosfato acumuladas. Este es el caso de Spirillum volutans.
Inclusiones de azufre
Las bacterias fotosintéticas (bacterias rojas y verdes) y cianobacterias utilizan el azufre y los sulfuros como fuente de poder reductor en la fotosíntesis (en bacterias con fotosíntesis anoxigénica). El exceso de azufre puede ser acumulado como inclusiones en el citoplasma, ya que el azufre libre es tóxico. Los gránulos se azufre se forman mientras están en el medio los compuestos reducidos del azufre. Cuando esto no están se oxidan.
Hay un grupo especial de bacterias, los quimiolitótrofos, las bacterias Beggiatoa/Thiothrix, que oxidan sustancias inorgánicas en sulfatos. También pueden acumular azufre en la célula.
Formaciones o corpúsculos intracelulares
En ocasiones, se les llama orgánulos, aunque no es muy correcto, ya que no hay orgánulos en procariotas.
Vacuolas de gas
Son corpúsculos presentes en bacterias acuáticas (de agua dulce o salada), en cianobacterias, y en una archaea, Halobacterium. Su misión es asegurar la flotabilidad de la célula en la columna de agua. Así, se sitúan en lugares donde la disponibilidad de oxigeno, la luz disponible y la concentración de nutrientes se adecue a sus necesidades. Están formadas por membranas compuestas únicamente de proteínas, sin lípidos, que se polimerizan formando un cilindro hueco que es impermeable al agua pero totalmente permeable a los gases de la atmosfera exterior, de tal manera que la composición de la vacuola dependerá de la composición de la atmosfera exterior.
Además de ayudar en la flotabilidad, las vacuolas de gas sirven para soportar la presión del medio.
Carboxisomas o cuerpos poliédricos
Aparecen en cianobacterias, en bacterias nitrificantes y, en general, en todas las bacterias que utilizan CO2 (como las bacterias reductoras de azufre). La fijación del CO2 ocurre dentro de estos carboxisomas que contienen una enzima clave: ribulosa-1,5-bifosfato-carboxilasa, mas conocida como rubisco. Están rodeados de una monocapa proteica a modo de cubierta aislante.
Clorosomas
Son las llamadas vesículas de Chlorobium (bacteria verde). Están muy próximas pero independientes de la membrana plasmática. Constan de una monocapa lipídica que las envuelve y les da forma cilíndrica, y contienen una gran parte del aparato fotosintético de las bacterias verdes. La bacterioclorofila (equivalente a la clorofila de plantas) está en la membrana, no en los clorosomas. No todas las bacterias fotosintéticas lo tienen.
Magnetosomas
Cristales o acúmulos de magnetita (Fe3O4). Para sintetizarse dependen de que en el medio haya siempre hierro. Si las cultivamos en un medio en ausencia de hierro, nunca se forman los magnetosomas y se pierden estas propiedades. Normalmente los magnetosomas se sintetizan por bacterias anaerobias estrictas o microaerófilas. Por ejemplo, Aquaspirillum tiene magnetosomas.
Endospora bacteriana
Es una estructura rígida, específica y muy diferenciada se que produce dentro de la célula. Ocurre en algunas especies bacterianas, principalmente bacilos (aunque no exclusivamente) que tienen el suelo como hábitat. Las endosporas son muy rígidas y muy resistentes al calor pero también a la desecación, a las radiaciones (tanto ionizantes como ultravioleta) y a ciertos compuestos de origen sintético que reciben el nombre de xenobióticos (ajenos a la naturaleza, como los antibióticos). Una vez formadas pueden permanecer viables durante larguísimos periodos de tiempo (se han recuperado algunas de hasta 5000 años). Con un buen microscopio se ven refringentes, pero su estructura rígida e impermeable hace que sean muy difíciles de teñir con colorantes, por lo que hay que aplicar técnicas especiales de tinción.
Las bacterias nunca forman endosporas cuando están creciendo, en fase exponencial. Requiere que haya, o bien limitación de algún nutriente esencial, o que haya cambios bruscos en las condiciones del medio. Mientras haya crecimiento activo nunca se forman endosporas. Para cada especie, la localización y tamaño de la endospora es específico. Pueden ser centrales, terminales o intermedias. La endospora puede permanecer viable mucho tiempo, y, si las condiciones son favorables, germinan y se vuelven a convertir en células funcionales.
Cuando hablamos de procariotas (y nos vamos a centrar casi exclusivamente en bacterias) las esporas se forman en el interior de la célula y se denominan endosporas. Sin embargo, en eucariotas y actinomicetos, las esporas se forman en el exterior de la célula, a veces dentro de estructuras especializadas, y se denominan exosporas. Eso quiere decir que en bacterias una célula solo puede generar una única espora (1 célula → una espora). En eucariotas y actinomicetos una célula puede formar un número indeterminado de esporas (1 célula → n esporas).
En bacterias, la endosporulación es un mecanismo de resistencia (al calor, la desecación, las radiaciones, etc). En eucariotas y actinomicetos, se forman esporas como mecanismo de reproducción y colonización de nuevos territorios (mecanismo de multiplicación). Por ello, el término espora en procariotas y eucariotas designa estructuras distintas.
Microorganismos que forman endosporas
En principio esta capacidad está restringida a unos pocos grupos de bacterias, los bacilos GRAM+, dentro del cual se encuentran los géneros Bacillus (aerobios estrictos) y Clostridium (anaerobios estrictos). También pueden formar endosporas algunos cocos, como el género Sporosarcina y Desulfotomaculum (GRAM-) (el resto son GRAM+).
Estructura de una endospora
La endospora está compuesta por varias capas:
- Primera capa: exosporio. Película muy fina formada por proteínas. No está presente en todas las endosporas.
- Segunda capa: cubierta de la endospora. Más densa, con grosor mayor, formada por proteínas ricas en aminoácidos con azufre (cisteína y metionina) que permiten formar enlaces covalentes a través de puentes disulfuro entre sus cadenas laterales. Es la capa responsable de la resistencia que tiene la endospora frente a las radiaciones y a la deshidratación.
- Tercera capa: córtex. Se corresponde con una capa muy laxa de peptidoglicano: posee un bajo nivel de tetrapéptidos anclados, y, en consecuencia, el nivel de transpeptidación es bajo también.
- Cuarta capa: pared celular. Se corresponde con la pared celular de la célula que se ha endosporulado y que va a servir como matriz cuando las condiciones del medio sean favorables y la endospora germine. Esta matriz va a servir para sintetizar pared nueva.
- Quinta capa: aparece el citoplasma rico en ribosomas.
- Sexta capa: nucleoide.
La endospora se encuentra en un estado quiescente, latente, de criptobiosis, aunque no es metabólicamente inerte. Tiene un cierto metabolismo basal.
El córtex y la cubierta no producen resistencia a la temperatura, solo a la desecación y radiación. La resistencia a la temperatura depende de un compuesto químico exclusivo: el ácido dipicolínico, que solo es sintetizado durante la formación de la endospora. Proviene de la ruta de biosíntesis de aminoácidos y puede llegar a representar un 10-20% del peso seco total de la endospora. El ácido dipicolínico se asocia con calcio y forma dipicolinato cálcico que es el compuesto responsable de la termorresistencia bacteriana. Es exclusivo de endosporas.
Etapas de la formación de una endospora
La endosporulación es un mecanismo de diferenciación celular mediante el cual una célula que llamamos vegetativa (una célula normal) se va a convertir en otra célula especial, termorresistente, y que puede llevar una vida latente indefinida. Así que, sí, las endosporas son células. En este proceso ocurren cambios estructurales, modificaciones genéticas (se van a expresar genes nuevos y se van a silenciar otros), cambios metabólicos, y va a ocurrir una deshidratación importante en lo que es la región central de la espora. Si la célula no sufre ningún tipo de limitación nutricional o de alteración ambiental nunca va a formar endosporas. Este proceso ocurre solo cuando aparecen las circunstancias adversas.
Primero, hay una replicación del nucleoide (material genético) en paralelo con la división celular. Las dos copias del nucleoide se unen entre sí formándose un doble filamento de ADN. En este momento ocurre un repliegue interno solo de la membrana plasmática, y captura o encierra una de las copias del ADN. Si la tabicación de la membrana plasmática se completa (se forma un septo), obtenemos una célula interna pequeñita dentro de la célula inicial: la pre-espora. Hasta este momento, el proceso es reversible. Si las condiciones mejoran, se puede revertir y no se forma la espora. De la pre-espora en adelante ya no se puede revertir.
A partir de aquí, la membrana completa el proceso de aislamiento hasta aislar a la espora dentro del citosol celular como una estructura independiente. Una vez que tenemos la espora individualizada se van a ir sintetizando las diferentes capas hasta formar la espora completa. En este momento, se libera al medio formando una espora libre. En este estado de endospora libre puede aguantar mucho tiempo. Cuando las condiciones mejoran, la endospora germina. La célula vegetativa que germina en el interior se desprende de las cubiertas de la endospora y sale al exterior. Esta germinación necesita de una fase previa de activación, generalmente por calor.
A nivel metabólico, ocurren varios procesos:
- Síntesis de dipicolinato cálcico.
- Enorme pérdida de agua: la endospora es una estructura muy deshidratada, con un 10-30% del agua de una célula vegetativa normal.
- Bajada de pH interno en aproximadamente una unidad (paralelo a la deshidratación). Esto, junto con la deshidratación, va a enlentecer la actividad de las enzimas y van a hacer que la endospora sea menos sensible a los radicales libres que podrían provocar radiaciones. También van a reducir su sensibilidad al efecto de las sustancias oxidantes.
- Síntesis de proteínas SASPs. Son proteínas específicas de la endosporulación que van a tener varias funciones: unirse y estabilizar el material genético (de tal manera que el ADN se hace mucho más resistente al calor y a la radiación), y servir como fuente de carbono y energía cuando se inicia la germinación de la endospora.
Cristales paraesporales de Bacillus Thuringiensis
La especie Bacillus Thuringiensis, cuando forma endosporas, sintetiza una estructura cristalina que es considerada como un insecticida biológico muy eficaz. Este cristal es tóxico, así que, cuando las larvas y orugas de los insectos (por ejemplo) lo ingieren se disuelve en sus jugos gástricos y se convierte en una toxina letal.
Actualmente, se usa como un insecticida biológico en los cultivos de cereales, frutas, verduras y hortalizas. El gen que codifica este cristal se ha clonado por modificación génica, y con ese gen se han obtenido plantas transformadas (tomate y algodón).


