Todas las proteobacterias quimioorganótrofas aerobias son GRAM negativas (-). Vamos a hablar de 3 grupos: Las Pseudomonadáceas, las bacterias del ácido acético y las fijadoras de nitrógeno.
Índice de contenidos
Pseudomonadáceas
El género conocido como Pseudomonas constituye un grupo heterogéneo que pertenece a 3 clases de protebacterias (α, β y γ). Se clasifican en 7 grupos atendiendo a homologías del rRNA, que se han subdividido según rasgos fenotípicos: presencia de PHβ, producción de un pigmento fluorescente, patogenicidad, presencia de arginina-dihidrolasa, empleo de glucosa, etc.
Son bacilos rectos o ligeramente curvados, móviles, con flagelación polar, y quimioheterótrofas. Son fáciles de cultivar porque no tienen requerimientos nutritivos complejos. Algunas se pueden considerar quimiolitoautótrofas como Pseudomonas pseudovorans, capaz de oxidar el hidrógeno. Son catalasa + y oxidasa +.
Llevan a cabo respiración aeróbica por la vía de Entner-Doudoroff (ED), con el ciclo de Krebs completo. Presentan oxidación completa de la glucosa, pudiendo usar como aceptor final de electrones oxígeno o nitratos (como Pseudomonas denitrificans).
Hay una serie de pruebas identificativas que ayudan a distinguir pseudomonadáceas de enterobacterias. Por ejemplo, la ausencia de producción de gas en el consumo de glucosa: se coloca un tubo pequeño con caldo de cultivo y se le introduce una campana de Durham (que es un tubo pequeño también) y ver si se forma una burbujita, lo que sería indicativo de la producción de gas. En pseudomonadáceas no se produce gas. La prueba O/F (oxidación+/fermentación-) permite distinguir los microorganismos que llevan a cabo la respiración oxidativa de la fermentación.
Muchas producen una serie de pigmentos hidrosolubles que excretan al medio. Pseudomonas aeruginosa produce un pigmento azul denominado piocianina, u otro verde en algunas cepas: pioverdina. Hay pseudomonas (como Pseudomonas fluorescens) que son fluorescentes a luz ultravioleta.
Podemos hallar pseudomonadáceas en aguas, suelos, y muchos otros hábitats. Hay inocuas y algunas son patógenas de animales como P. aeruginosa o de plantas, como Xanthomonas campestris. Muchas especies son mesófilas, pero algunas son psicrófilas. Son las responsables de proporcionar sabores, olores, colores, a ciertos alimentos refrigerados en mal estado (producen cambios en las propiedades organolépticas de los alimentos)
Pueden usar como fuente de carbono y energía compuestos pocos corrientes, incluso algunos de origen sintético: xenobióticos, como pesticidas y otros productos químicos tóxicos, gracias a la producción de una serie de enzimas capaces de degradar estos compuestos. Estos enzimas están codificados por plásmidos. Desde el punto de vista medioambiental les permite ser considerados como agentes de biorremediación (uso de organismos para tratar de eliminar/solucionar problemas medioambientales, como contaminación).
Aunque algunas especies pueden causar auténticos problemas en lugares como hospitales o laboratorios de síntesis de medicamentos y otras sustancias, ya que son capaces de crecer sobre adhesivos, jabones o antisépticos (compuestos de carbono cuaternario).
Suelen producir bacteriocinas (proteínas de elevado peso molecular que afectan a especies cercanas a las especies que las producen). En el caso concreto de pseudomonadáceas se denominan biocinas.
Pseudomonas aeruginosa
Es la especie más significativa. Es un patógeno oportunista, especialmente en hospitales. Las infecciones adquiridas en hospitales se denominan infecciones nosocomiales. No causa problemas en individuos sanos, pero si el individuo está inmunocomprometido (defensas bajas), puede llegar a infectar heridas, quemaduras, tracto urinario; y causar meningitis, abscesos e incluso septicemia (enfermedad asociada con la presencia en sangre de agentes patógenos o toxinas bacterianas producidas por esos patógenos).
Es resistente a la mayoría de los antibióticos de uso común, aunque hay algunos a los que es sensible: esta resistencia está relacionada por una parte con el pequeño tamaño de las porinas y a la presencia de genes que confieren resistencia, localizados en los plásmidos R.
Zooglea
Posee la típica morfología de bacilo con flagelación polar como Pseudomonas aeruginosa. Es importante para el tratamiento aerobio de aguas residuales, ya que oxida con mucha eficacia grandes cantidades de materia orgánica del efluente hasta CO2 y agua.
Xanthomonas
Es fitopatógeno. Causa varias lesiones necróticas en plantas. Se caracteriza por el tipo de pigmentos que produce: amarillos, muy llamativos. Es de importancia comercial, incluye una especie que se cultiva porque su cápsula contiene goma xantano (X. campestris), que se usa como espesante en muchas áreas industriales (farmacéutica, agroquímica, cosmética, alimenticia...).
Está siendo usada en programas de recuperación secundaria de petróleo, ya que es capaz de reducir la permeabilidad y movilidad del agua donde se ha producido un vertido de petróleo al incrementar su viscosidad.
Bacterias del ácido acético
Todas son α-proteobacterias. Son bacilos GRAM negativos (-), aerobios y móviles. Pueden llevar a cabo la oxidación incompleta de alcoholes y azúcares, lo que provoca la acumulación de ciertos ácidos orgánicos como producto final (como el ácido acético si usan etanol). Presentan una tolerancia relativamente alta a los ácidos (la mayoría puede crecer a pH inferior a 5) lo cual es esencial para organismos que produzcan ácidos.
Son un grupo heterogéneo que incluye organismos con flagelación polar o perítrica. Los que tienen flagelación polar se clasifican dentro del género Gluconobacter, los que tienen flagelación perítrica están englobados en el género Acetobacter. Se diferencian en su capacidad de oxidar posteriormente el acético para generar CO2. Si tienen el ciclo de Krebs completo, como Acetobacter, pueden oxidar completamente el acético. Acetobacter oxida hasta CO2 + H2O mientras que le quede etanol.
Etanol → Acético → CO2 + H2O
O sea, el último paso solo lo puede hacer cuando ya no queda nada o muy poco de etanol.
Gluconobacter tiene el ciclo de Krebs incompleto y no lo pueden realizar (no lo puede oxidar hasta CO2 y agua). Mientras quede alcohol no usan el ácido acético.
Se encuentran en bebidas alcohólicas y se aíslan a menudo de la sidra, el vino o la cerveza. En placas con carbonato cálcico (CaCO3) y etanol se diferencian fácilmente: el acético producido disuelve el carbonato cálcico del medio. Al disolverse se produce un aclaramiento en la placa alrededor de las colonias que hayan producido ácido acético.
Se emplean en la producción de vinagre.
Además de usar el etanol pueden oxidar de manera incompleta otros compuestos orgánicos como alcoholes de cadena larga y azúcares como glucosa. Ésta es oxidada solo hasta glucónico, y la galactosa hasta galactónico. Esta característica suboxidante es aprovechada para la producción del ácido ascórbico (Vitamina C). Ésta es fácil de producir a partir de la sorbosa, y estas bacterias son capaces de convertir el sorbitol hasta sorbosa. Es un proceso de biotransformación.
Otra propiedad bastante interesante es su capacidad de sintetizar celulosa. Esta celulosa no difiere sustancialmente de la de origen vegetal, salvo que es pura y no está mezclada con otros polímeros como hemicelulosa, pectinas o ligninas. No forma parte de la pared celular, sino que forma una matriz externa por fuera de la pared (se observan rodeadas de una maraña de microfibrillas de celulosa). Puede usarse como espesante en alimentos. Cuando se cultivan en medio líquido se crea una película de celulosa. Esta maraña puede suponer una estrategia para estar siempre en contacto con el oxígeno en la superficie de líquido.
Bacterias fijadoras de nitrógeno
Diversos organismos del suelo son fijadores aeróbicos del nitrógeno. Están en 3 clases: α, β o γ proteobacterias.
Azotobacter
Son γ-proteobacterias (gamma), fijadores libres de nitrógeno. Tiene forma de bacilo pleomórfico (forma siempre definida) de gran tamaño (cercano al de las levaduras), de más de 2 a 4 μm, más bien cortos. Son aerobios estrictos y fijan nitrógeno de forma no simbiótica. Algunas cepas son móviles gracias a flagelación perítrica y otras cepas son inmóviles.
Cuando fijan nitrógeno en un medio con carbohidratos, producen unas cápsulas mucosas/mucilaginosas. Realizan respiración aerobia. Su nitrogenasa es sensible al oxígeno, pero representan la tasa respiratoria más elevada de todos los microorganismos: esto está relacionado con la protección de la nitrogenasa, ya que el oxígeno que entra al interior celular rápidamente se reduce a agua y de esta forma se evita que se inactive la nitrogenasa, además de la ayuda de la cápsula mucosa. Además, la nitrogenasa tiene protección conformacional: forma asociaciones con una proteína determinada que la inactiva en presencia de oxígeno.
Pueden formar cistes o quistes. Son formas de resistencia, pero no de latencia como las endosporas. Presentan bajos niveles de respiración endógena, son resistentes a la desecación, rotura mecánica, radiación ultravioleta e ionizante, pero no son particularmente resistentes al calor. No están completamente latentes, si se les suministra una fuente de carbono utilizable, la oxidan rápidamente.
Rhizobium
Son α-proteobacterias aeróbicas. Incluye bacilos móviles con flagelación polar o perítrica según la especie y con un tamaño medio de 0'5-0'9 x 1'2-3 μm. Presentan pleomorfismo en condiciones adversas (cambian de forma).
Crecen en simbiosis en el interior de las células de los nódulos radiculares de las leguminosas. Son muy específicos, una cepa bacteriana puede establecer simbiosis con una determinada especie de leguminosa, pero no con otras. No pueden fijar nitrógeno por separado, necesitan las condiciones microaerófilas que le aporta la planta.
En el nódulo radicular debe de haber unos niveles bajos de oxígeno, que están controlados por la leghemoglobina, una ferroproteína roja que se une al oxígeno. La formación de esta leghemoglobina esta inducida por la interacción de los dos organismos: la parte globina está formada por la planta, y el grupo hemo esta codificado por la bacteria. Podemos considerar que la leghemoglobina actúa a modo de tampón de oxígeno, manteniendo bajo y constante el nivel de oxígeno próximo a la bacteria (condición indispensable para que fije nitrógeno).
Fijan nitrógeno en estado de bacteroide en los nódulos radiculares de leguminosas. El bacteroide así se convierte en microaerófilo.
Formación del nódulo
Reconocimiento y adherencia
El contacto entre la bacteria y la raíz se lleva a cabo por parte de los dos organismos. La planta produce unas lectinas (glicoproteínas que son capaces de aglutinar a las bacterias). Se han identificado tanto en los extremos de los pelos radiculares como en la superficie de las bacterias próximas a las raíces.
Por otra parte, la bacteria lleva a cabo la unión por una porción polisacarídica de la membrana externa de su pared (parte polisacárida del lipopolisacárido). Presentan también unos pili que participan en la unión a los pelos radiculares.
También actúan unas proteínas especiales de unión denominadas ricadhesinas, que se unen a calcio y captan complejos de calcio en la superficie de los pelos radiculares.
Enrrollamiento e invaginación de los pelos radiculares
Debido a unas sustancias denomianas factores Nod (codificadas por los genes Nod, producidos por la bacteria, pero solo en presencia de los flavonoides producidos por la planta) se va a producir el enrollamiento del pelo radicular. Son sustancias producidas por las bacterias, pero inducidas por flavonoides liberados por la raíz de la planta.
Entrada en los pelos radiculares
Las bacterias entran porque inducen la formación por parte de la planta de canales de infección en los pelos radiculares, por donde pasan a través las bacterias.
Invasión y desplazamiento
Las bacterias van avanzando a través de los canales de infección hasta alcanzar células adyacentes a los pelos radiculares. Normalmente éstas son células diploides, y, si la alcanzan, sufren necrosis y degeneran. Pero también pueden alcanzar células tetraploides, activando su división gracias a los factores Nod (producidos por la bacteria). Esto desencadena la formación del nódulo.
O sea, si infectan células diploides, estas van a degenerar y morir, sufriendo necrosis. Pero si infectan células tetraploides se desencadena la formación de nódulos.
Formación de nódulos
Las bacterias comienzan a multiplicarse en el interior de las células tetraploides y sufren unas modificaciones hasta convertirse en bacteroides: formas ramificadas, hinchadas y deformes, que se rodean de forma individual o en pequeños grupos de la membrana celular de la planta. La membrana que rodea a los bacteroides se denomina peribacteroide. Sólo se fija nitrógeno cuando se han formado los bacteroides y se han rodeado de la membrana peribacteroide. Se denomina entonces simbiosoma (bacteroide + peribacteroide), que pueden ser individuales o estar dispuestos en pequeños grupos.
Cuando la planta muere, los nódulos se deterioran y parte de esas bacterias pasan al suelo. Cuando las bacterias se encuentran en estado de bacteroide no pueden dividirse, pero siempre dentro de esos grupos habrá alguna bacteria que no se habrá convertido a bacteroide y estará de forma latente. Estas sí que pueden proliferar en el suelo e iniciar la infección de otras raíces cercanas.
Fijación bioquímica del nitrógeno
La fijación se realiza desde nitrógeno atmosférico hasta amoníaco.
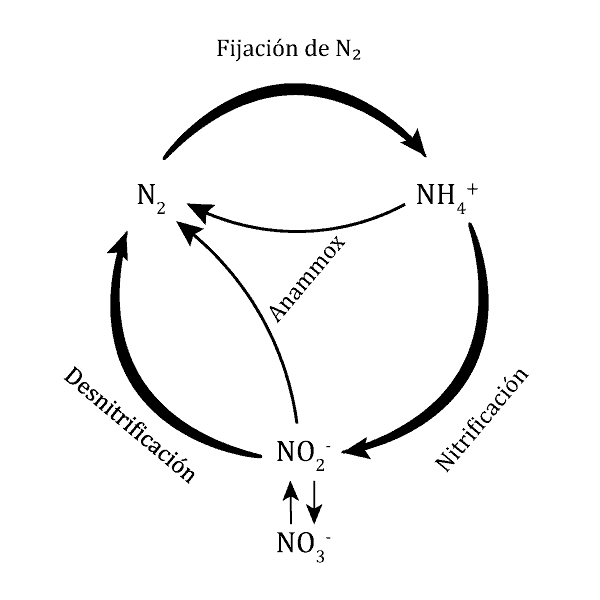
En este caso, la reacción se realiza única y exclusivamente en el simbiosoma. Las nitrogenasas se encuentran siempre en el interior del bacteroide. El bacteroide es totalmente dependiente de la planta para fijar nitrógeno, ya que ésta reduce la presión parcial de oxígeno: la leghemoglobina se combina con el oxígeno para evitar que entre en contacto con la nitrogenasa. La planta le suministra nutrientes, principalmente intermediarios del ciclo TCA (succinato, malato, fumarato) los cuales serán donadores de electrones en última instancia para la obtención de ATP y para la propia reacción de la nitrogenasa (que requiere ATP + poder reductor).
La bacteria le suministra el nitrógeno asimilado. El primer producto estable es el amoníaco que puede formar parte de aminoácidos o se puede convertir en glutamina por la glutamina-sintasa: la concentración de glutamina-sintasa en el interior del bacteroide es baja, pero en la célula vegetal adyacente es elevada, por lo que en la planta pasará rápidamente de amoníaco a glutamina. La planta sintetiza amidas, ureidos, a partir de la incorporación del amoníaco.
Factores Nod
Los genes responsables de la formación del nódulo en la bacteria, los genes Nod, se encuentran en unos plásmidos de gran tamaño denominados plásmidos Sym. En ocasiones están rodeados de genes Nif donde está codificada la nitrogenasa.
En algunos casos lo genes Nif se encuentran en el cromosoma, no en el plásmido. Algunos de los plásmidos Sym son responsables de la especificad de las cepas bacterianas para establecer simbiosis con determinadas plantas de leguminosas, formando los denominados grupos de inoculación cruzada, que son cepas de Rhizobium que pueden infectar a un grupo de especies próximas de leguminosas. Las transferencias de estos plásmidos permiten que cepas que no puedan infectar ahora sí lo puedan hacer.
Los factores Nod, dependiendo de las especies tienen variaciones en las cadenas laterales.