
Otras bacterias (volumen V)
En el volumen V del Manual de Bergey se engloban 10 filos que no encajan en los grupos anteriores. Todas son GRAM negativas (-), pero existe gran variabilidad de morfología, fisiología y estilos de vida.
Índice de contenidos
Planctomyces
Bacteria acuática que tiene forma globular y posee un pedúnculo largo y estrecho al final del cual hay un disco adhesivo. A diferencia de otras pedunculadas, su pedúnculo no consiste en una extrusión citoplasmática con pared celular, sino que es de naturaleza proteica y se le califica como tallo. Sirve para adherirse a superficies o entre ellas.
Es una bacteria gemante con un ciclo de vida similar a las bacterias pedunculadas: con una fase nadadora y otra pedunculada. La forma nadadora se origina por gemación del cuerpo celular; se une a una superficie y comienza a formar un pedúnculo, que se va a ir alargando y, llegado un punto, genera una nueva célula por gemación.

.
Es quimioheterótrofa y anaerobia facultativa. Esta bacteria y sus relacionadas ocupan una posición filogenética específica, de forma independiente al resto. Carecen de peptidoglicano; su pared celular es proteica, de tipo capa S, rica en cisteína y prolina. Al no tener peptidoglicano, son resistentes a los antibióticos que actúan sobre este, como la penicilina. Por ello, para facilitar su aislamiento, se añade penicilina al medio.
Los planctomicetos poseen una compartimentalización interna, muy desarrollada, que incluye algo parecido a una estructura nuclear. Ciertos compartimentos carecen de DNA, y tienen otras funciones. Algunos parientes de Planctomyces son Gemmata y Brocadia anammoxidans.
El compartimento interno más grande es la membrana intracitoplasmática (ICM), separada de la membrana citoplasmática por una región libre de ribosomas denominada parifoplasma. Después de la ICM está el riboplasma. En el caso de Gemmata el nucleoide está dentro de una membrana del cuerpo nuclear; en Brocadia no ocurre esto, el compartimento más grande es el anamoxosoma, donde se realiza la reacción de anamox, y el DNA está fuera de la membrana del cuerpo nuclear.
Clamidias
Son parásitas intracelulares estrictas, es decir, deben crecer y multiplicarse dentro de células hospedadoras. Probablemente representan un paso más allá en la evolución en la pérdida de independencia; tienen menos funciones metabólicas propias que las rickettsias. Desarrollan una estrategia muy efectiva de supervivencia que incluye el parasitismo de los recursos del hospedador y la producción de una forma de resistencia para su transmisión. Están relacionadas con multitud de enfermedades y de síndromes.
Durante mucho tiempo fueron consideradas como formas intermedias entre los virus y las bacterias, por ser parásitos intracelulares, pero en realidad son bacterias ya que tienen DNA y RNA. Se pueden considerar las formas más simples de organización celular. Cuando entran en una célula hospedadora no pierden su integridad estructural, a diferencia de los virus. Son inmóviles, pequeñas y con un genoma de tan solo 4-6 x 108 Da.

Tipos de células
Tienen un ciclo vital que implica dos tipos de células: el cuerpo elemental (EB), que es la forma infecciosa, y el cuerpo reticulado (RB), que es la forma vegetativa.
El cuerpo elemental tiene forma de coco y no se multiplica, posee un material genómico electrón-denso y una pared celular rígida. Es relativamente resistente a ciertas condiciones adversas (desecación) y es la forma especializada en la dispersión; se transmite por el aire sin necesidad de vectores.
En cuanto al cuerpo reticulado, es la forma intracelular especializada en la división celular por fisión binaria. No es infecciosa. Tiene una forma algo más alargada y más grande que el cuerpo elemental. Posee un material genómico menos denso y contiene más ribosomas que el cuerpo elemental; y su pared es más flexible.
Aunque la envoltura de las clamidias se asemeja a la de otras GRAM negativas (-), no poseen peptidoglicano ni amino-azúcares semejantes. Poseen una capa P, proteica. Los EB alcanzan su estabilidad osmótica mediante enlaces cruzados entre proteínas de su membrana externa (CRP: proteínas ricas en cisteína), estableciendo puentes disulfuro entre estas cisteínas. También hay enlaces entre las proteínas periplasmáticas. Estos puentes disulfuro se reducen químicamente y son causa de la transformación de los cuerpos elementales a los reticulados.
Ciclo vital
Comienza con la fijación de un EB a la superficie de una célula hospedadora, que lo fagocita por endocitosis en lo que será el cuerpo de inclusión. La clamidia permanece dentro del endosoma y se reorganiza a cuerpo reticulado. Algún componente del cuerpo elemental inhibe la fusión del fagosoma al lisosoma; además, se producen cambios a nivel de la membrana del cuerpo de inclusión, permitiéndose la entrada de nutrientes del hospedador hacia la clamidia.
El cuerpo elemental se va transformando en cuerpo reticulado ya que va aumentando de tamaño, perdiendo rigidez de su pared, e inicia la síntesis de macromoléculas. Es probable que la síntesis inicial de RNA y proteínas vaya destinada a la producción de más ribosomas. Transcurridas de 8 a 10 horas, los cuerpos reticulados inician la fisión binaria, y tras 24-30 horas comienzan a transformarse en cuerpos elementales. Continúan el proceso hasta que la célula hospedadora se lisa (como en C. psittaci) y libera las clamidias o sale por un proceso de endocitosis inversa (como en C. pneumoniae).
Metabolismo
Su capacidad biosintética está más restringida que en el caso de rickettsias. De hecho, durante mucho tiempo se pensó que eran parásitos energéticos estrictos (necesitan robar ATP). No obstante, se ha comprobado, mediante la secuenciación de su genoma, que presentan genes implicados en la síntesis del ATP. No generan ATP porque poseen una translocasa de membrana del mismo tipo que el de las rickettsias, que pueden intercambiar ADP endógeno por ATP exógeno. Entonces, podemos decir que no son "parásitos energéticos estrictos".
Incluso, aunque carecen de peptidoglicano, presentan genes implicados en la síntesis de éste. Son sensibles a algunos antibióticos que inhiben la síntesis del peptidoglicano (ejemplo de pérdida evolutiva de función).
Cuando el hospedador aporta los precursores, los cuerpos reticulados pueden sintetizar sus propios componentes. Poseen porinas y proteínas de transporte activo de membrana, aunque se conoce poco de este transporte. Los cuerpos elementales poseen una actividad metabólica muy reducida, ya que están especializados en la dispersión.
Constituyen una rama independiente del dominio bacteria, aunque algo próxima a los planctomicetales. Hay dos géneros: Chlamydia y Chlamydophila.
Chlamydia
Desarrolla un solo cuerpo de inclusión, consecuencia de la fusión de los distintos endosomas formados por los diferentes cuerpos elementales que entran a la célula. En este cuerpo, Chlamydia retiene glucógeno como fuente de energía, y tiene un único operón ribosomal.
C. trachomatis es la especie más estudiada; posee muchos serotipos agrupados en 3 biotipos principales:
- Causante del tracoma. Infección crónica de la córnea y la conjuntiva y es el principal motivo de ceguera en el hombre. Causa dolor e inflamación.
- Causante del linfogranuloma venéreo: mucho más invasivo que el anterior, es una ETS. Causa también la uretritis no gonocócica. Afecta al tracto genitourinario, podemos calificarlo como el principal causante de las ETS más frecuentes.
- Afecta al ratón.
Chlamydophila
No acumula glucógeno y tiene 2 operones ribosomales.
- C. psittaci causa la psitacosis tanto en hombres como animales, considerada como una enfermedad epizoótica en aves, aunque también afecta a gatos, vacas, ovejas, etc. No sólo se transmite por contacto con las heces de los psitácidos, sino también de otras aves. Pude provocar diferentes tipos de infecciones gastrointestinales, afectación a la placenta, ojos, etc. Se contagia a través de la inhalación de los cuerpos elementales.
- C. pneumoniae puede provocar diferentes síndromes respiratorios en el hombre.
Espiroquetas
Las bacterias del filo Spirochaetes son gram negativas (-), quimioheterótrofas y poseen una morfología helicoidal, más o menos laxa dependiendo del género y especie. Son bastante delgadas y largas. Poseen el cuerpo cilíndrico, constituido por el protoplasma, y, a parte de la membrana, está rodeado por pared celular y una vaina (membrana externa), con 3 capas. El cuerpo cilíndrico es rígido, mientras que la vaina externa es flexible. La vaina rodea tanto al periplasma, la pared celular como al cuerpo cilíndrico. En el periplasma están los filamentos axiales.
El filamento axial rodea la espiroqueta y le permite el movimiento; está formado por fibrillas axiales conocidas como endoflagelos o flagelos periplásmicos, en número de 2 a más de 100, y están entre el peptidoglicano y la vaina (membrana externa). Se fijan cerca de los extremos celulares y se extienden hasta aproximadamente dos tercios de la longitud celular. Son rígidos y rotan unidos a cada extremo celular.
Como el cilindro protoplasmático también es rígido, al girar provocan en la vaina una onda o cresta helicoidal de tal manera que el cilindro protoplasmático gira en la dirección opuesta. Si está en contacto con alguna superficie el movimiento es de reptación. Si se daña o se elimina la vaina o filamento axial, la espiroqueta ya no puede moverse. Usan básicamente el mismo mecanismo de movimiento que otras bacterias, la rotación de flagelos rígidos adheridos a la membrana citoplasmática.
Están bien adaptadas a los medios viscosos que dificultan o impiden el movimiento por flagelos típicos. Esto explica por qué varias especies parásitas están restringidas a tejidos y fluidos animales.
Hay mucha variedad en cuanto a requerimiento de oxígeno: aerobias, anaerobias y anaerobias facultativas. Todas se dividen por fisión binaria.
El hábitat también es muy variado, aparecen tanto en ambientes acuáticos como terrestres, son frecuentes en lodos con materia orgánica en descomposición. Muchas son patógenas de hombre y animales.
Familia Spirochaetaceae
Spirochaeta
Es una bacteria acuática, inocua, de vida libre. Unas son anaerobias y otras son anaerobias facultativas. Junto con Cristispira es de tamaño muy superior a las demás, del orden de 2 a 40 filamentos axiales. Aparece en aguas dulces y marinas.
Cristispira
Es también acuática e inocua. Se considera comensal en el tracto digestivo de moluscos, aunque no se ha podido cultivar. Aparece exclusivamente en el estilo cristalino de moluscos, aunque no en todas las especies de moluscos. No se conoce la razón fisiológica de que aparezca en ese hábitat ni hay pruebas de que resulte nociva para su hospedador. Aparece más frecuentemente en moluscos sanos que enfermos. Tiene más de 100 fibrillas axiales en el filamento axial.
Treponema
Anaerobio con algunas especies microaerófilas. Hay especies comensales y otras patógenas: T. oralis y T. denticola son típicas de la boca, T. saccharophilum está presente en el rumen de rumiantes. T. pallidum causa la sífilis en el hombre. La vaina es muy delgada y son más pequeñas que las anteriores.
Borrelia
Es un género patógeno que suele usar vectores artrópodos. Tiene un cromosoma lineal y es de tamaño similar a Treponema.
B. recurrentis causa las fiebres recurrentes, que, si no tienen tratamiento pueden causar la muerte. Es una enfermedad sistémica debilitante que se transmite por un piojo.
B. burgdorferi causa la enfermedad de LYME, extendida en unas zonas de EEUU con una garrapata específica.
Familia Leptospiraceae
Son todas aerobias.
Leptospira
Es el género representativo, con dos especies.
- Dentro de Leptospira biflexa hay del orden de 60 serotipos y es de vida libre.
- L. interrogans es parásita patógena, con 218 serotipos establecidos, aunque que no todos afectan al hombre. El serotipo canicola no afecta al hombre, pero icterohaemorragiae sí. La enfermedad de Weil es causada en el hombre por el serotipo icterohaemorrhagiae, y a veces por el canicola. Esta enfermedad es una zoonosis que en ocasiones afecta al hombre. Se multiplica en diferentes partes del cuerpo y luego se pasa al hígado y riñones produciendo diversos daños, y puede contaminar por orina. Es de crecimiento lento. Utiliza como fuente de carbono ácidos grasos y alcoholes.
Acidobacterias
Forman parte del Phylum Acidobacteria. Es un grupo diverso (25 subgrupos) e importante por su amplia distribución en ambientes acuáticos y terrestres. Son las bacterias más abundantes del suelo.
Sólo se han podido cultivar unas pocas; de las restantes no se conoce tanto, sólo por estudios moleculares del SSU rRNA. Son muy abundantes en cuevas, del orden del 25% de la población de organismos presentes, además de ser las más abundantes de suelos ácidos.
De lo anterior podemos deducir que sí, son acidófilas. Algunas que son capaces de emplear el FeS están implicadas en la decoloración de objetos y obras de arte, así como de pinturas rupestres. Son GRAM negativos (-), quimioheterótrofas, de crecimiento muy lento. Tienen capacidades metabólicas variadas, unas respiran y otras fermentan; e incluso ejercen la fotosíntesis, como Chloroacidobacterium, que hace fotosíntesis anoxigénica. Es difícil inferir su papel en los ambientes donde se encuentran al ser tan abundantes.
Una de las que se ha podido cultivar es Acidobacterium, la cual es acidófila, aerobia y capsulada. Se ha aislado de aguas ácidas de drenaje minero. Usa tanto azúcares como ácidos orgánicos; algunas especies son anaerobias.
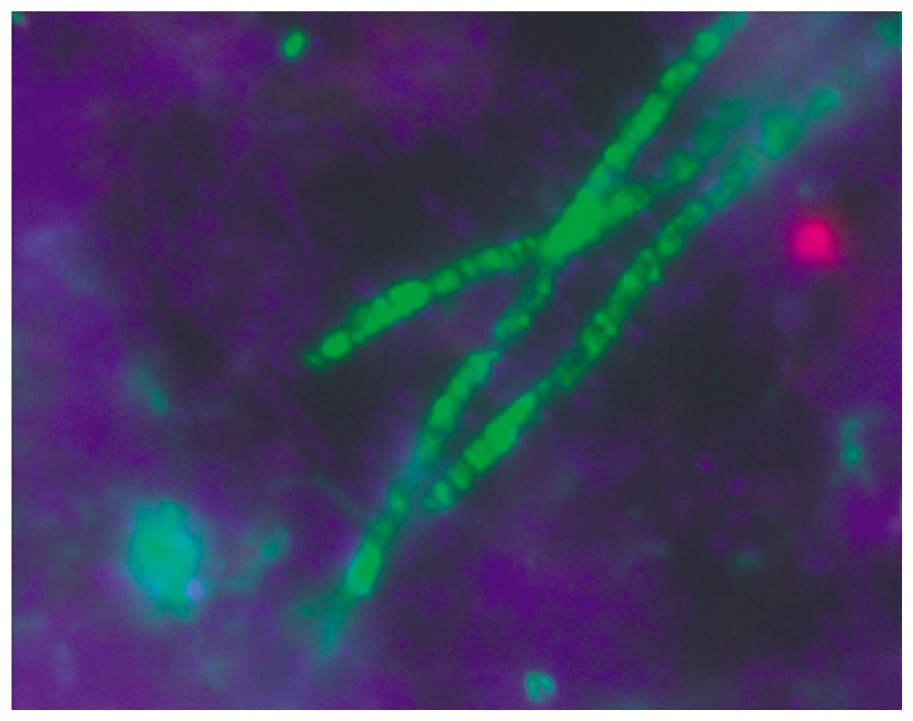
Bacteroidetes
El filo Bacteroidetes cuenta es bacilo gram negativo (-) anaerobio estricto, no formador de endosporas, cuyo género más representativo es Bacteroides. Es quimioheterótrofo, y fermenta azúcares y proteínas, y como producto se obtiene alguna mezcla de ácidos orgánicos (acetato y succinato).
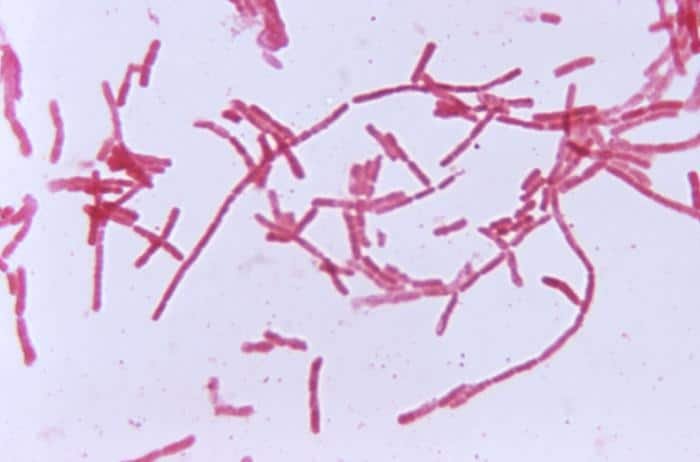
Son normalmente comensales en el tracto intestinal humano y de otros animales.
B. ruminicola aparece en el rumen de rumiantes. Son las más abundantes del intestino delgado del hombre. Algunas son patógenas; las anaerobias son las más importantes que están asociadas con infecciones humanas que pueden afectar desde el sistema nervioso central al sistema esquelético.
B. fragilis es muy frecuente en infecciones abdominales, pélvicas, pulmonares y sanguíneas.
Grupo Cytophagales
Son bacilos muy delgados, largos, GRAM(-) y a menudo, con los extremos puntiagudos, que se desplazan por deslizamiento. Están ampliamente distribuidas en agua y suelos donde pueden ser muy abundantes. Se enmarcan dentro del filo Bacteroidetes.
Sporocytophaga forma unas estructuras de latencia esféricas denominadas microcistos, similares a las bacterias nitrificantes. Junto con Cytophaga son aerobias, responsables de gran parte de la degradación bacteriana de celulosa en ambientes aerobios naturales.
Muchas degradan polisacáridos pero no mediante la secreción de enzimas extracelulares, sino por enzimas ligados a la pared celular. Esto explica por qué necesitan un contacto directo con las fibrillas de celulosa para degradarlas. También degradan quitina, queratina, pectina; incluso algunas marinas degradan el agar.
Desempeñan un papel muy importante en la mineralización de la materia orgánica y causan daños en aparejos de pesca y estructuras de madera expuestas. Aparecen en plantas de tratamiento de aguas residuales. Algunas son patógenas de peces como C. columnaris, que puede causar serios problemas en piscifactorías.
El movimiento parece que se debe a unas partículas esféricas de naturaleza proteica que rotan en el periplasma, a expensas del potencial de membrana o por gasto directo de ATP. Esto permite el deslizamiento de la célula a lo largo de las superficies.
Tiene una serie de ventajas:
- Hace que la bacteria entre en contacto con las fuentes de nutrientes insolubles, como celulosa o quitina
- Está bien adaptado a hábitats más secos y al movimiento dentro de masas sólidas como tierra, sedimentos, o madera podrida.
- Permite ubicar a la bacteria en las condiciones ambientales óptimas de densidad de luz, oxígeno, temperatura, etc.