El phylum Actinobacteria (actinobacterias o actinomicetos) es extenso y complejo: está formado por 1 clase, 6 órdenes y 44 familias taxonómicas. Son bacterias GRAM+ con alto contenido en G+C.
Son actinomicetos (filamentosos, frecuentemente ramificados) y parientes con un alto contenido G+C (>50% mol). Respecto a su morfología, podemos decir que suelen ser bacilos o filamentos principalmente aerobios. Su hábitat son suelos y material vegetal.
En su mayoría son comensales inofensivos, aunque Corynebacterium, Nocardia y Mycobacterium contienen importantes patógenos para los seres humanos. Hay un interés economico en estos, ya que algunos producen antibióticos, o productos lácteos fermentados.
Índice de contenidos
Micrococcus
Se agrupan en cocos (parejas, tétradas o agrupaciones irregulares) y tienen alto contenido en G+C. Son gram positivos (+) y catalasa (+). Suelen ser inmóviles. Siempre son cocos, y sus colonias están pigmentados por carotenoides (amarillo, naranja o rojo). Están ampliamente extendidos en el suelo, el agua y sobre la piel de mamíferos.
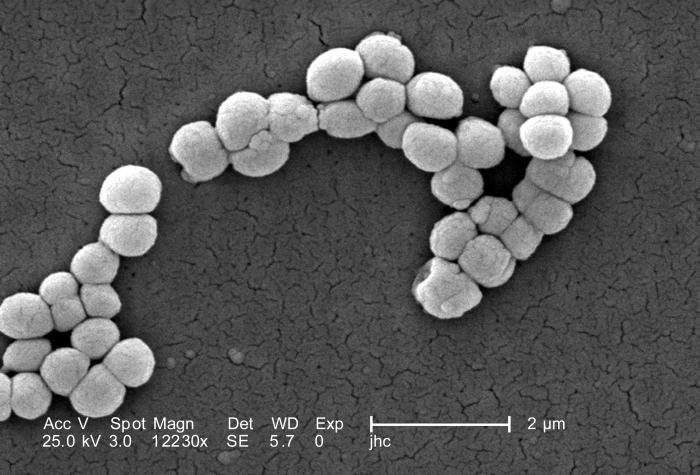
Corinebacterias
También denominadas bacterias corinefoides. Son bacilos móviles con un crecimiento con forma irregular (letras chinas, mazo, forma de V) que se debe a una división crepitante/angular/por fractura que produce un movimiento de desgarro cuando se dividen (la membrana tiene 2 capas). Ambas bacterias quedan unidas por la pared externa y el septo que las separa es la pared interna. Después, se rompe la capa externa en un lado sólo y quedan como una bisagra.
Corinebacterium
Este género es anaerobio facultativo siempre catalasa +. Respira O2 (respiración aerobia) y la fermentación es propiónica, sin O2. Presenta ácidos micólicos (3-hidroxiácidos ramificados) pero no son ácido-alcohol resistentes (tinción Ziehl-Neelsen).
Respecto a las especies principales de Corinebacterium podemos destacar:
- Especies patógenas. La más importante es C. diphtheriae (bacilo de Klebs-Löeffler), aunque también están C. bovis, C. equi y C. betae
- Otras especies son inocuas como:
- C. pseudodiphtheriticum. Se encuentra en el tracto respiratorio.
- C. xerosis. Aparece en la boca y piel; además, forma parte de la microbiota normal.
- C. glutamicum. Se encarga de la producción de ácido glutámico que se emplea como aditivo alimenticio, de gran importancia industrial.
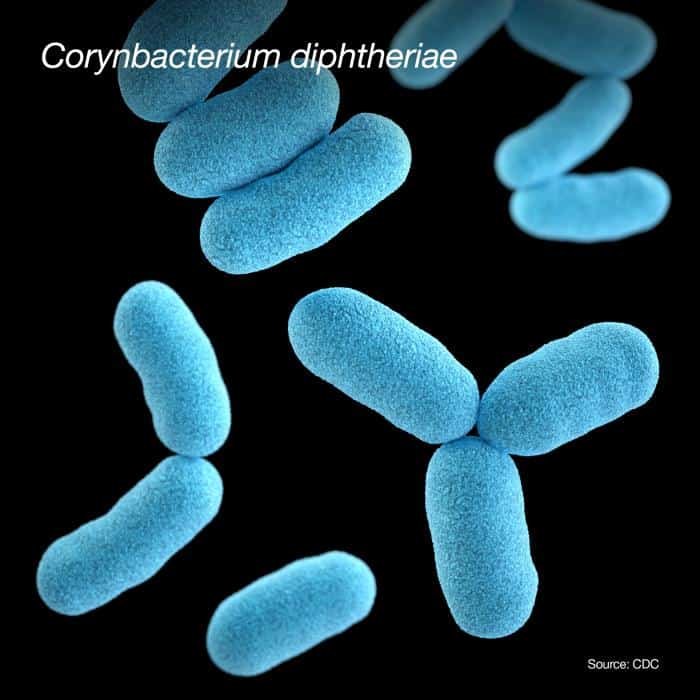
Corinebacterium diphtheriae
Es la principal causa de difteria. Se utiliza agar telúrito (agar cistina-TeO3K) para su aislamiento. Este agar inhibe el crecimiento de cualquier muestra nasofaringial. Forma colonias negras en agar telúrito y blancas en agar-sangre.
La toxicidad de esta bacteria es por la exotoxina diftérica. La toxina la codifica el fago β que contiene el gen tox. Hay conversión lisogénica (genes del fago alteran el fenotipo de la bacteria hospedadora). Las cepas no lisogénicas son atóxicas, no producen la enfermedad. Por otro lado, las cepas virulentas son así por la presencia del fago en estado lisogénico.
La expresión del gen tox depende del estado fisiológico de la bacteria. La presencia de hierro es un condicionante. El gen se expresa cuando los niveles de hierro son bajos, debido a que su alta concentración inhibe la expresión del gen.
La exotoxina entra en las células e inhibe el EF2 de las células (inhibe la traducción) impidiendo la unión del aminoácido a la cadena de aminoácidos nacientes. No se consiguen formar proteínas por lo que la célula muere. Cuando se produce la exotoxina ésta pasa al torrente sanguíneo pero la bacteria se queda en la boca y provoca la pseudomembrana diftérica por falta de O₂.
Existen 3 biotipos:
- gravis: exotoxina más potente. En agar aparece gris y rugosa.
- intermedius: colonias pequeñas y puntiformes grises.
- mitis: menos potente. Colonias negras, brillantes y convexas.
Arthrobacter
Es aerobio estricto, por lo que en la pruebas es catalasa +. Presenta dimorfismo. En la fase exponencial se muestra como un bacilo. Tiene división crepitante formando ramificaciones irregulares de aspecto poco filamentoso. También presenta fragmentación múltiple. En fase estacionaria, tiene forma de cocos que se diferenciarán hasta bacilos.
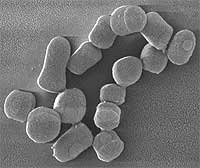
Es un importante agente mineralizador de la materia orgánica en el suelo; muy resistente a la desecación y a la privación de nutrientes. Está adaptado a este medio. Tiene una extraordinaria flexibilidad nutricional poco común al degradar pesticidas, algo muy inusual.
Propionibacterias
Propionibacterium
Tiene una morfología similar a corinebacterias (es un género muy pleomórfico). Puede denominarse corinebacteria anaerobia. Es anaerobio aerotolerante: siempre presenta fermentación propiónica (tiene pigmentos hémicos, citocromos, catalasa, etc). Es prácticamente anaerobio en cuestiones de O₂ ya que vive en lugares con presión parcial de O₂ baja. Tiene cadena de transporte de electrones (CTE) por lo que sería más correcto denominarlo anaerobio facultativo.
Su hábitat son los ambientes con bajas presiones parciales de O₂: tracto intestinal, genital, rumen y piel (Propionibacterium acnes), además de productos lácteos. De hecho, la primera vez que se aisló fue a partir de un queso suizo.
Fermenta azúcares y produce propionato, acetato, CO₂ y ± succinato. Los productos de su fermentación son utilizados por otros microorganismos para realizar la fermentación láctica. Puede fermentar lactato (fermentación secundaria).
Ciclo del propionato:
1 glucosa → 2 piruvato → 2 propionato.
A partir de lactato:
3 lactato → 3 piruvato → 2 propionato + 1 acetato + 1 CO₂
Formación de succinato:
fosfoenolpiruvato (PEP) + CO₂ → oxalacetato →→→ succinato
Propionibacterium acnes
Es causante del acné. Hay una producción alta de sebo por la actuación de hormonas y la presencia del microorganismo que segrega lipasas que lo degradan. Los productos generan inflamación al atravesar la dermis. Actualmente es denominado Cutibacterium acnes.

Bifidobacterium
Es un género diferente: son bacilos inmóviles aislados, o en parejas en forma de V o grupos, con extremos espatulados, y en muchos casos bifurcados.
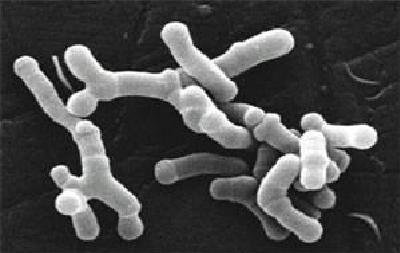
Son anaerobios estrictos o anaerobios aerotolerantes (según la especie): soportan O₂ siempre que haya CO₂.
Bifidobacterium bifidum
Es una fracción mayoritaria de la microbiota intestinal natural del recién nacido alimentado con leche materna. En el adulto, se encuentra en el intestino y en la boca. También aparece en otras especies, aguas residuales e insectos.
Necesita de aminoazúcares presentes en la leche (N-acetilglucosamina y N-acetilgalactosamina); si le faltan estos aminoazúcares cambian su morfología creando ramificaciones que dan lugar a un micelio rudimentario. El metabolismo de estas bacterias es similar al de las fermentadoras del ácido láctico. El lactato formado a partir de los aminoazúcares no se forma igual que en las fermentadoras del ácido láctico (realizan una fermentación via pentosas-fosfato).
2 glucosa → 3 acetato + 2 lactato (5 ATP) (no suelen producir CO2)
Micobacterias (Mycobacterium)
Son bacilos inmóviles aerobios, y presentan crecimiento filamentoso transitorio (fase estacionaria) que se rompe cuando llegan a la fase exponencial.
Tienen un alto contenido en lípidos en su pared (AAR, ácido-alcohol resistentes). Los lípidos son ácidos micólicos, lípidos de 60-90 carbonos con otros en posición 3-OH que dan lugar a los 3-hidroxiácidos grasos ramificados. Los ácidos micólicos les confieren un aspecto denso, compacto, céreo y rugoso a las colonias en medio sólido. En medio líquido forman grumos. Además, reducen la permeabilidad, aumentan la resistencia a la desecación y a agentes microbianos.
La presencia de ácidos micólicos es una propiedad extraña para el resto de bacterias excepto para bacterias no cardiformes y micobacterias. El tamaño de los ácidos es proporcional a la resistencia. También tienen una importante aplicación clínica en la detección de especies patógenas.
Clasificación
Velocidad de crecimiento
- De crecimiento rápido y no patógenas: Mycobacterium phlei, M. fortuitum y M. smegmatis.
- De creciemiento lento y patógenas de:
- hombre; Mycobacterium leprae (enfermedad de Hansen) y M. tuberculosis (bacilo de Koch).
- animales, y que pueden afectar al hombre; Mycobacterium avium subespecie paratuberculosis.
Pigmentación (carotenoides)
- No pigmentadas: Mycobacterium tuberculosis.
- Producen pigmentos en presencia de luz (fotocromogénesis).
- Producen pigmentos incluso en la oscuridad (escotocromogénesis).
Mycobacterium tuberculosis
Presenta crecimiento lento y es patógena del hombre. Es resistente a los germicidas y a pH alcalinos (aislamiento selectivo). Con estas características se pueden preparar medios selectivos.
Existe una correlación con la creación de formas de cordón al generarse por la agregación de estas bacterias (que forman cadenas alargadas) con la presencia de glicolípido; factor cuerda, responsable de esta formación, y de su virulencia (inhibe el paso de unión del fagosoma con el lisosoma).
Actinobacterias filamentosas
Son quimioheterótrofos y Gram positivos (+). Respecto a su morfología podemos decir que tienen un crecimiento ramificado similar al de los hongos filamentosos.
- Tienen un micelio no fragmentable y son capaces de formar esporas unicelulares asexuales
- Su desarrollo micelial es transitorio solo en crecimiento activo y fragmentable al reducirse la velocidad de crecimiento
- Tienen micelio de sustrato (hifas rizoides) y aéreo (hifas aéreas). Si producen esporas se sitúan en el extremo de las hifas aéreas. Si realizamos una sección transversal, en el centro hay esporas maduras en el micelio; en los extremos casi no presenta y en su superficie tampoco.
Método reproductivo
El método reproductivo es asexual por crecimiento apical de hifas. Se forman tabiques en el micelio que dan lugar a las esporas: conidiosporas (conidios) o esporangiosporas. Ambas son exosporas asexuales.
- Conidios: cuando las esporas no se encuentran encerradas en una estructura.
- Esporangios: cuando se encuentran encerradas en una estructura.
Su disposición y morfología tiene carácter sistemático. Se desarrollan por privación de nutrientes. No soportan las altas temperaturas, pero sí la desecación.
Metabolismo
La mayoría son inmóviles. Las esporas se transportan por viento o animales. Cuando hay movimiento, éste se limita a las esporas que presentan flagelos.
La pared celular es muy variada (60 unidades de peptidoglicano (PG); respecto al tercer aminoácido que ocupa lugar en la cadena de aminoácidos lo hace por puente intrapeptídico). El tipo de pared celular es un criterio taxonómico, que los divide en 4 grupos.
El porcentaje de G+C que presentan estas bacterias es del 63-78%. Tienen el contenido más alto en G+C. Se les considera un grupo muy coherente.
Viven libres, en el suelo, donde degradan grandes cantidades y variedades de compuestos orgánicos. Son esenciales en la mineralización de la materia orgánica. Además, se encuentran en la piel, y algunos pueden ser patógenos para el hombre, otros animales y algunas plantas.
Producen metabolitos secundarios con gran interés práctico: son los mayores productores de antibióticos junto con Bacillus y hongos eucariotas. Producen también fármacos anticancerígenos, antihelmínticos e inmunosupresores (útiles en procesos de transplante de órganos).
Nocardia
Cuenta con un micelio aéreo limitado (a partir de él pueden producir conidios) y es de sustrato fragmentable. Es una bacteria aerobia, catalasa +. Además, tiene la capacidad de usar una gran variedad de hidrocarburos, azúcares y ceras.
En la pared celular tiene peptidoglicano con mDAP y sin puente peptídico de unión. Posee ácidos micólicos (AAR).
Son saprófitos de vida libre, en suelos y aguas (deterioro de juntas de goma en tuberías de agua).
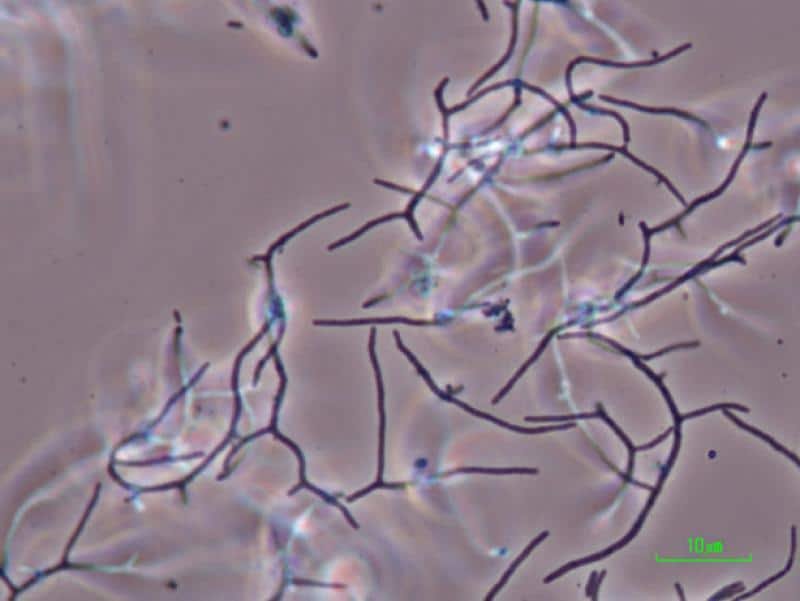
Nocardia asteroides. Es una bacteria patógena oportunista que afecta al hombre y otros animales en diferentes órganos. Pueden centrarse en alguno, como por ejemplo en pulmón, causando la nocardiosis pulmonar crónica que es difícil de tratar.
Streptomyces
Cuenta con más de 500 especies. Tienen un %GC de entre 69-78%. Forman colonias compactas. Las especies se establecen mediante características morfológicas y fisiológicas. Son aerobios estrictos.
Cuentan con micelio aéreo y de sustrato (no fragmentable). Sus filamentos son de 0,5-1µm Ø y longitud indefinida, y a menudo, sin septos en fase vegetativa.
Forman conidiosporas inmóviles en el extremo de las hifas aéreas (3 → 50) formadas a partir de esporóforos multinucleados, y hay una posterior separación de las células individuales.

La forma, textura y disposición de las esporas se utiliza como carácter taxonómico. Hay pigmentación de esporas y esporóforos. A veces, el micelio de sustrato también está pigmentado.
Este género es de fácil identificación en placa: tiene aspecto pulverulento, naturaleza compacta y color. Presentan enzimas hidrolíticos que degradan gran cantidad de compuestos. Son muy versátiles y adaptables. Cuentan con hidrolasas extracelulares de compuestos inusuales.
Esporulan sobre sustratos sólidos (agar) o en la superficie de cultivos líquidos sin agitación. Sólo colonizan medio sólido.
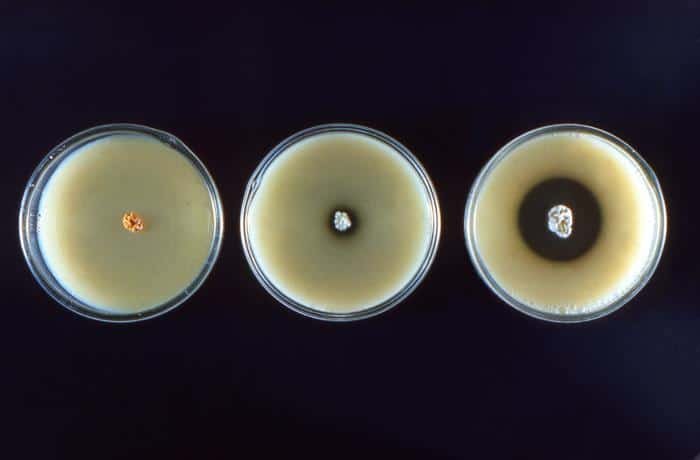
Su hábitat es principalmente de suelos (alcalinos y neutros), y son de fácil aislamiento Producen geosminas. A excepción de las especies que se citarán a continuación, el resto son inocuas:
- Streptomyces scabies: enfermedades en patata y remolacha.
- S. somaliensis: provoca un actinomicetoma.
- S. albus: dolencias y causas de patologías. Este microorganismo produce geosminas (olor terroso cuando llueve).
Streptomyces y los antibióticos
El género Streptomyces fabrica muchísimos antibióticos (±50%); cuenta con más de 10000 compuestos diferentes con actividad biológica. Producen ±2/3 partes de los agentes antimicrobianos empleados en medicina y veterinaria.
Algunos sintetizan más de 1 antibiótico, y a menudo, sin relación química. El mismo antibiótico puede ser producido por diferentes especies muy alejadas entre sí. Un cambio nutricional puede causar un cambio en la naturaleza del antibiótico. La bacteria productora resiste a su propio antibiótico pero puede ser sensible a otros.
Hay especies de Streptomyces con plásmidos lineales (500kb de longitud), implicados en la biosíntesis de antibióticos.
La búsqueda de nuevos antibióticos prosigue: el desarrollo de cepas resistentes requiere el descubrimiento continuo de nuevos agentes. Se ha usado la tecnología informática para encontrar nuevos antibióticos a partir de especies que ya conocemos; además de comprobar otras alternativas usadas en el pasado, como la terapia con bacteriófagos.
No se saben las razones ecológicas por las cuales llevan a cabo la producción de antibióticos. Se producen en fase estacionaria, por lo que no intervienen en el crecimiento, así que se cree que promueven la esporulación para incrementar sus posibilidades de supervivencia.
En el siguiente recuadro se pueden ver una serie de especies de Streptomyces con su correspondiente antibiótico generado y su diana celular.
| Microorganismo | Antibiótico | Estructura química | Diana celular |
|---|---|---|---|
| S. griseus | Estreptomicina | Aminoglucósido | 30S |
| S. orientalis | Vancomicina | Carboxiglicósido | Pared celular |
| S. venezuelae | Cloranfenicol | Derivado del benceno | 50S |
| S. mediterranei | Rifampicina | Ansamicina | RNA-pol del DNA |
| S. fradiae | Neomicina | Aminoglucósido | 30S |
| S. kanamyceticus | Kanamicina | Aminoglucósido | 30S |
| S. erythraeus | Eritromicina | Macrólido | 50S |
| S. aureofaciens | Tetraciclina | Tetraciclina | 30S |
Uno de los grandes desafíos actuales que afronta la ciencia es la aparición de bacterias resistentes a los antibióticos. Esto supone la necesidad de descubrir, o bien nuevos antibióticos, o bien otros métodos de eliminación de dichas bacterias. Y, aunque este desafío recae especialmente en la medicina y la microbiología, hay otras ramas científicas que podrían ayudar a la resolución de este problema, como por ejemplo, las ciencias de la computación; concretamente la inteligencia artificial.
Un grupo de investigadores ha conseguido entrenar a una red neuronal para que tenga la capacidad de predecir moléculas con capacidad antibacteriana. Para ello, han realizado predicciones en diversas bibliotecas químicas, y se ha descubierto un compuesto con la capacidad antibacteriana deseada, la halicina.
Índice de contenidos
El problema de los antibióticos: búsqueda de soluciones
Este hallazgo llega en el mejor momento, ya que desde hace décadas no se descubre ningún nuevo antibiótico; solo aparecen nuevas generaciones de antibióticos ya existentes. Esto, junto con la aparición cada vez más común de bacterias resistentes a los antibióticos, hace que la necesidad de encontrar nuevos antibióticos sea urgente. Se han están probando otras alternativas, como renovar el uso bacteriófagos en terapia antimicrobiana. Pero otra opción es encontrar nuevos antibióticos con ayuda de la computación.
No es nuevo el intento usar la informática para encontrar nuevos antibióticos. Ya en el pasado se han desarrollado modelos para predicciones de las propiedades de ciertas moléculas. No obstante, la falta de precisión de estos modelos hacia que estos intentos fueran en vano. Ahora bien, los nuevos avances de los algoritmos de los modelos neuronales hacen que aparezca una oportunidad para descubrir nuevos medicamentos. Ahora, la red neuronal es capaz de predecir las propiedades de una molécula de manera precisa, si bien todavía hay diferencias entre los enfoques analítico y experimental. Entonces, la mejor solución sería unir ambos enfoques.
Halicina: resultado de unir los enfoques analítico y experimental
Esto es lo que hicieron este grupo de investigadores. Primero, entrenaron a un modelo de red neuronal para predecir la inhibición del crecimiento de Escherichia coli usando una colección de 2.335 moléculas. Después, aplicaron este modelo a un conjunto de librerías químicas que englobaban más 107 millones de moléculas, para identificar compuestos con potencial actividad contra E. coli. Por último, tras clasificar a dichos compuestos moleculares en función de la puntuación otorgada por el modelo de predicción, se seleccionaron una lista de candidatos viables.
Mediante este método, identificaron un compuesto de una librería química llamada 'Drug Repository Hub', concretamente el inhibidor SU3327 de la kinasa c-jun N-teminal, renombrado como halicina. Este compuesto, diferente a nivel estructural a los antibióticos convencionales, se mostró como un potente inhibidor del crecimiento de E. coli.
Del mismo modo, la halicina muestra actividad bactericida frente a un amplio espectro de bacterias infecciosas, que incluyen a Mycobacterium tuberculosis (bacteria de la tuberculosis) y Enterobacterias resistentes a carbapenem. Además, también ha demostrado su efectividad contra Clostridioides difficile y Acinetobacter baumannii pan-resistentes en modelos murinos (modelo de ratones para investigación). Esta propiedad antibacteriana es debido a que interrumpe el flujo de protones a través de la membrana celular. Además, en los experimentos mostró una baja toxicidad y dificultad a la aparición de resistencias.
Además de la halicina, se han identificado ocho componentes antibacterianos que son estructuralmente diferentes de los antibióticos conocidos. De esas moléculas, dos mostraron actividad antibiótica de amplio espectro, y podrían tener la capacidad de vencer a cepas de E. coli resistentes a antibióticos.
Conclusión
Los modelos neuronales no son perfectos; hay que diseñar el entrenamiento de manera precisa para nuestro objetivo, se deben elegir bien las librerías químicas, y se debe tener claro cuales son las prioridades en las posibles predicciones. Aun con sus limitaciones, el uso de modelos de redes neuronales tiene el potencial de encontrar nuevos compuestos antibióticos con menos recursos, abaratando el coste de las investigaciones. Este no deja de ser un estudio preliminar, pero nos indica un posible camino hacia el descubrimiento de nuevos antibióticos, y la resolución del problema de las bacterias resistentes a éstos.
Bibliografía
Jonathan M. Stokes et al. A deep learning approach to antibiotic discovery. Cell, vol. 180, n.º 4, págs. 688-702 (2020). DOI: https://doi.org/10.1016/j.cell.2020.01.021