Saltar al contenido principal
Saltar al pie de página
Microbiología
Historia de la Microbiología
La célula procariota
Técnicas microbiológicas básicas
Nutrición microbiana. Medios de Cultivo
Microscopios, colorantes y tinciones
Tinciones y Colorantes principales en microbiología
Crecimiento y control
Análisis del crecimiento microbiano
Factores ambientales para el crecimiento microbiano
Métodos de Control del Crecimiento Microbiano
Agentes Quimioterápicos: los Antibióticos
Metabolismo Microbiano
Fototrofía. Fotosíntesis Oxigénica y Anoxigénica
Quimiolitotrofía
Quimioorganotrofía. Fermentación, Respiración Aerobia
Quimioorganotrofía. Respiración anaerobia
Fijación del nitrógeno en la naturaleza
Análisis necesarios para clasificación de procariotas
Bacterias
Clasificación de las bacterias
Volumen I
Arqueas
Aquifex y el grupo Deinococcus-Thermus
Bacterias fotosintéticas
Volumen II. Proteobacterias
Proteobacterias quimiolitótrofas y metanótrofas
Proteobacterias quimiorganótrofas aerobias
Proteobacterias quimioorganótrofas anaerobias facultativas
Rickettsias: otras Proteobacterias (I)
Otras Proteobacterias (II)
Volumen III. Firmicutes
Volumen IV. Actinobacterias
Volumen V. Otras bacterias
Eucariotas
Protistas
Hongos
Virus
Clasificación de los virus
Clasificacion ICTV (Nueva)
Clasificación de Baltimore
Virus Animales. Priones
Virus Vegetales. Viroides
Bacteriófagos: todo sobre esta posible alternativa a los antibióticos
Herramientas
Calculadora de curva de crecimiento bacteriano
Generador de citas bibliográficas
Glosario de microbiología
Blog
Microbiología
Historia de la Microbiología
La célula procariota
Técnicas microbiológicas básicas
Nutrición microbiana. Medios de Cultivo
Microscopios, colorantes y tinciones
Tinciones y Colorantes principales en microbiología
Crecimiento y control
Análisis del crecimiento microbiano
Factores ambientales para el crecimiento microbiano
Métodos de Control del Crecimiento Microbiano
Agentes Quimioterápicos: los Antibióticos
Metabolismo Microbiano
Fototrofía. Fotosíntesis Oxigénica y Anoxigénica
Quimiolitotrofía
Quimioorganotrofía. Fermentación, Respiración Aerobia
Quimioorganotrofía. Respiración anaerobia
Fijación del nitrógeno en la naturaleza
Análisis necesarios para clasificación de procariotas
Bacterias
Clasificación de las bacterias
Volumen I
Arqueas
Aquifex y el grupo Deinococcus-Thermus
Bacterias fotosintéticas
Volumen II. Proteobacterias
Proteobacterias quimiolitótrofas y metanótrofas
Proteobacterias quimiorganótrofas aerobias
Proteobacterias quimioorganótrofas anaerobias facultativas
Rickettsias: otras Proteobacterias (I)
Otras Proteobacterias (II)
Volumen III. Firmicutes
Volumen IV. Actinobacterias
Volumen V. Otras bacterias
Eucariotas
Protistas
Hongos
Virus
Clasificación de los virus
Clasificacion ICTV (Nueva)
Clasificación de Baltimore
Virus Animales. Priones
Virus Vegetales. Viroides
Bacteriófagos: todo sobre esta posible alternativa a los antibióticos
Herramientas
Calculadora de curva de crecimiento bacteriano
Generador de citas bibliográficas
Glosario de microbiología
Blog
Buscar
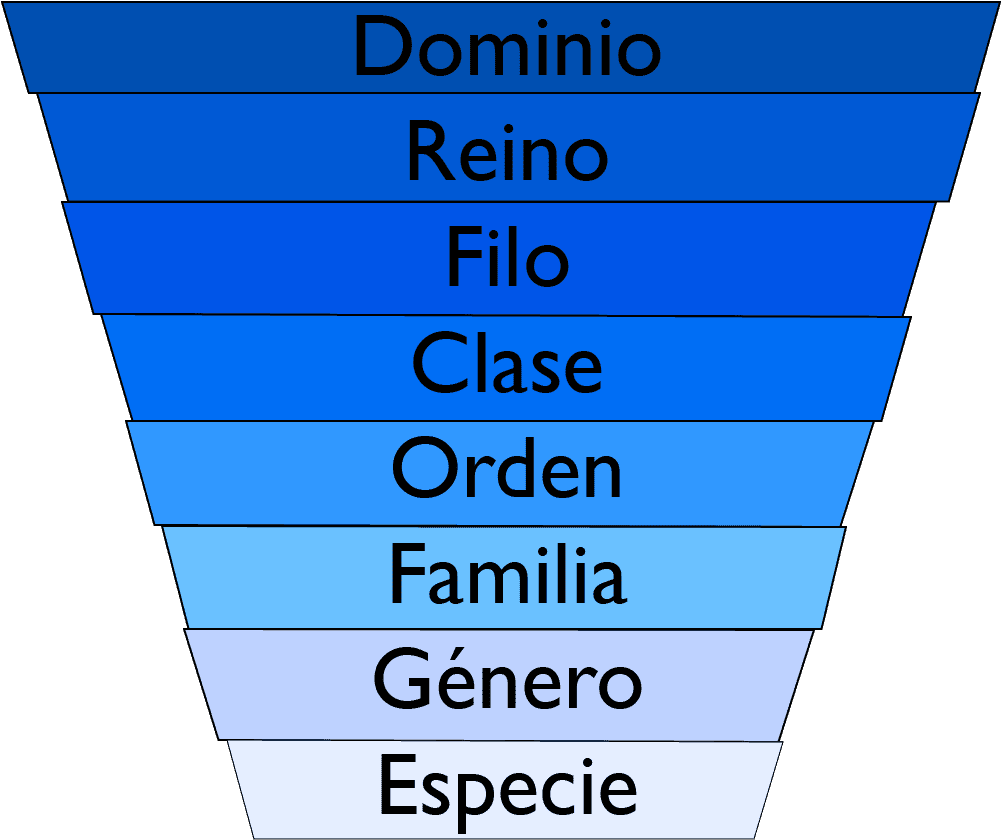
Información acerca de taxonomía en nuestro blog
Análisis necesarios para clasificación de procariotas
Clasificación de los seres vivos: taxonomía