Información acerca de virus en nuestro blog

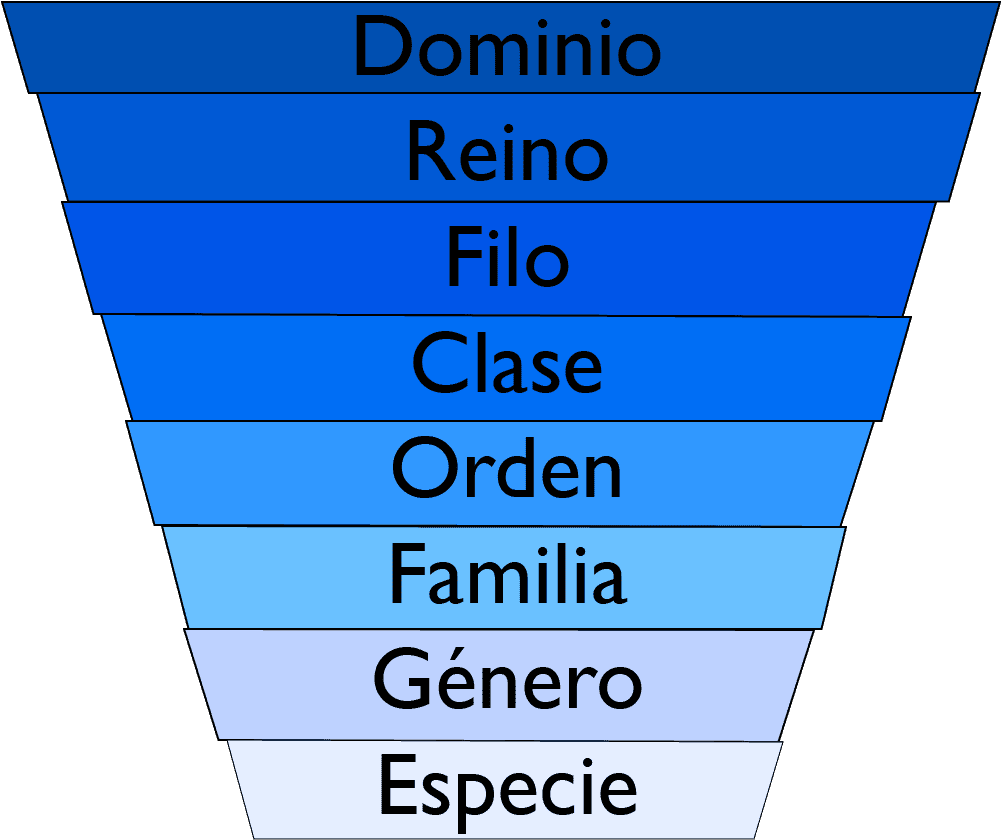
Clasificación de los seres vivos: taxonomía

¿Los asintomáticos no contagian? Screening en Wuhan
Clasificacion ICTV (Nueva)

Familia Filoviridae

Diagnosis de COVID-19

Factores que determinan el Ritmo de Propagación del Virus

¿Por qué es el Jabón es tan ÚTIL frente a los Virus?

Familia Hepeviridae
Clasificación ICTV

Clasificación de Baltimore

Virus Vegetales. Viroides
Clasificación de los virus

Virus: Caracteres generales

Bacteriófagos: todo sobre esta posible alternativa a los antibióticos
Coronavirus SARS-CoV-2 (COVID-19)
Familia Coronaviridae